WO1999025821A1 - Compositions and methods for genetic modification of plants - Google Patents
Compositions and methods for genetic modification of plants Download PDFInfo
- Publication number
- WO1999025821A1 WO1999025821A1 PCT/US1998/024610 US9824610W WO9925821A1 WO 1999025821 A1 WO1999025821 A1 WO 1999025821A1 US 9824610 W US9824610 W US 9824610W WO 9925821 A1 WO9925821 A1 WO 9925821A1
- Authority
- WO
- WIPO (PCT)
- Prior art keywords
- frt
- sites
- plant
- site
- recombinase
- Prior art date
Links
Classifications
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12N—MICROORGANISMS OR ENZYMES; COMPOSITIONS THEREOF; PROPAGATING, PRESERVING, OR MAINTAINING MICROORGANISMS; MUTATION OR GENETIC ENGINEERING; CULTURE MEDIA
- C12N15/00—Mutation or genetic engineering; DNA or RNA concerning genetic engineering, vectors, e.g. plasmids, or their isolation, preparation or purification; Use of hosts therefor
- C12N15/09—Recombinant DNA-technology
- C12N15/63—Introduction of foreign genetic material using vectors; Vectors; Use of hosts therefor; Regulation of expression
- C12N15/79—Vectors or expression systems specially adapted for eukaryotic hosts
- C12N15/82—Vectors or expression systems specially adapted for eukaryotic hosts for plant cells, e.g. plant artificial chromosomes (PACs)
- C12N15/8216—Methods for controlling, regulating or enhancing expression of transgenes in plant cells
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12N—MICROORGANISMS OR ENZYMES; COMPOSITIONS THEREOF; PROPAGATING, PRESERVING, OR MAINTAINING MICROORGANISMS; MUTATION OR GENETIC ENGINEERING; CULTURE MEDIA
- C12N15/00—Mutation or genetic engineering; DNA or RNA concerning genetic engineering, vectors, e.g. plasmids, or their isolation, preparation or purification; Use of hosts therefor
- C12N15/09—Recombinant DNA-technology
- C12N15/10—Processes for the isolation, preparation or purification of DNA or RNA
- C12N15/102—Mutagenizing nucleic acids
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12N—MICROORGANISMS OR ENZYMES; COMPOSITIONS THEREOF; PROPAGATING, PRESERVING, OR MAINTAINING MICROORGANISMS; MUTATION OR GENETIC ENGINEERING; CULTURE MEDIA
- C12N15/00—Mutation or genetic engineering; DNA or RNA concerning genetic engineering, vectors, e.g. plasmids, or their isolation, preparation or purification; Use of hosts therefor
- C12N15/09—Recombinant DNA-technology
- C12N15/11—DNA or RNA fragments; Modified forms thereof; Non-coding nucleic acids having a biological activity
- C12N15/113—Non-coding nucleic acids modulating the expression of genes, e.g. antisense oligonucleotides; Antisense DNA or RNA; Triplex- forming oligonucleotides; Catalytic nucleic acids, e.g. ribozymes; Nucleic acids used in co-suppression or gene silencing
- C12N15/1137—Non-coding nucleic acids modulating the expression of genes, e.g. antisense oligonucleotides; Antisense DNA or RNA; Triplex- forming oligonucleotides; Catalytic nucleic acids, e.g. ribozymes; Nucleic acids used in co-suppression or gene silencing against enzymes
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12N—MICROORGANISMS OR ENZYMES; COMPOSITIONS THEREOF; PROPAGATING, PRESERVING, OR MAINTAINING MICROORGANISMS; MUTATION OR GENETIC ENGINEERING; CULTURE MEDIA
- C12N15/00—Mutation or genetic engineering; DNA or RNA concerning genetic engineering, vectors, e.g. plasmids, or their isolation, preparation or purification; Use of hosts therefor
- C12N15/09—Recombinant DNA-technology
- C12N15/63—Introduction of foreign genetic material using vectors; Vectors; Use of hosts therefor; Regulation of expression
- C12N15/79—Vectors or expression systems specially adapted for eukaryotic hosts
- C12N15/82—Vectors or expression systems specially adapted for eukaryotic hosts for plant cells, e.g. plant artificial chromosomes (PACs)
- C12N15/8201—Methods for introducing genetic material into plant cells, e.g. DNA, RNA, stable or transient incorporation, tissue culture methods adapted for transformation
- C12N15/8202—Methods for introducing genetic material into plant cells, e.g. DNA, RNA, stable or transient incorporation, tissue culture methods adapted for transformation by biological means, e.g. cell mediated or natural vector
- C12N15/8203—Virus mediated transformation
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12N—MICROORGANISMS OR ENZYMES; COMPOSITIONS THEREOF; PROPAGATING, PRESERVING, OR MAINTAINING MICROORGANISMS; MUTATION OR GENETIC ENGINEERING; CULTURE MEDIA
- C12N15/00—Mutation or genetic engineering; DNA or RNA concerning genetic engineering, vectors, e.g. plasmids, or their isolation, preparation or purification; Use of hosts therefor
- C12N15/09—Recombinant DNA-technology
- C12N15/63—Introduction of foreign genetic material using vectors; Vectors; Use of hosts therefor; Regulation of expression
- C12N15/79—Vectors or expression systems specially adapted for eukaryotic hosts
- C12N15/82—Vectors or expression systems specially adapted for eukaryotic hosts for plant cells, e.g. plant artificial chromosomes (PACs)
- C12N15/8201—Methods for introducing genetic material into plant cells, e.g. DNA, RNA, stable or transient incorporation, tissue culture methods adapted for transformation
- C12N15/8202—Methods for introducing genetic material into plant cells, e.g. DNA, RNA, stable or transient incorporation, tissue culture methods adapted for transformation by biological means, e.g. cell mediated or natural vector
- C12N15/8205—Agrobacterium mediated transformation
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12N—MICROORGANISMS OR ENZYMES; COMPOSITIONS THEREOF; PROPAGATING, PRESERVING, OR MAINTAINING MICROORGANISMS; MUTATION OR GENETIC ENGINEERING; CULTURE MEDIA
- C12N15/00—Mutation or genetic engineering; DNA or RNA concerning genetic engineering, vectors, e.g. plasmids, or their isolation, preparation or purification; Use of hosts therefor
- C12N15/09—Recombinant DNA-technology
- C12N15/63—Introduction of foreign genetic material using vectors; Vectors; Use of hosts therefor; Regulation of expression
- C12N15/79—Vectors or expression systems specially adapted for eukaryotic hosts
- C12N15/82—Vectors or expression systems specially adapted for eukaryotic hosts for plant cells, e.g. plant artificial chromosomes (PACs)
- C12N15/8201—Methods for introducing genetic material into plant cells, e.g. DNA, RNA, stable or transient incorporation, tissue culture methods adapted for transformation
- C12N15/8213—Targeted insertion of genes into the plant genome by homologous recombination
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12N—MICROORGANISMS OR ENZYMES; COMPOSITIONS THEREOF; PROPAGATING, PRESERVING, OR MAINTAINING MICROORGANISMS; MUTATION OR GENETIC ENGINEERING; CULTURE MEDIA
- C12N15/00—Mutation or genetic engineering; DNA or RNA concerning genetic engineering, vectors, e.g. plasmids, or their isolation, preparation or purification; Use of hosts therefor
- C12N15/09—Recombinant DNA-technology
- C12N15/63—Introduction of foreign genetic material using vectors; Vectors; Use of hosts therefor; Regulation of expression
- C12N15/79—Vectors or expression systems specially adapted for eukaryotic hosts
- C12N15/82—Vectors or expression systems specially adapted for eukaryotic hosts for plant cells, e.g. plant artificial chromosomes (PACs)
- C12N15/8241—Phenotypically and genetically modified plants via recombinant DNA technology
- C12N15/8261—Phenotypically and genetically modified plants via recombinant DNA technology with agronomic (input) traits, e.g. crop yield
- C12N15/8271—Phenotypically and genetically modified plants via recombinant DNA technology with agronomic (input) traits, e.g. crop yield for stress resistance, e.g. heavy metal resistance
- C12N15/8274—Phenotypically and genetically modified plants via recombinant DNA technology with agronomic (input) traits, e.g. crop yield for stress resistance, e.g. heavy metal resistance for herbicide resistance
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12N—MICROORGANISMS OR ENZYMES; COMPOSITIONS THEREOF; PROPAGATING, PRESERVING, OR MAINTAINING MICROORGANISMS; MUTATION OR GENETIC ENGINEERING; CULTURE MEDIA
- C12N15/00—Mutation or genetic engineering; DNA or RNA concerning genetic engineering, vectors, e.g. plasmids, or their isolation, preparation or purification; Use of hosts therefor
- C12N15/09—Recombinant DNA-technology
- C12N15/87—Introduction of foreign genetic material using processes not otherwise provided for, e.g. co-transformation
- C12N15/90—Stable introduction of foreign DNA into chromosome
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12N—MICROORGANISMS OR ENZYMES; COMPOSITIONS THEREOF; PROPAGATING, PRESERVING, OR MAINTAINING MICROORGANISMS; MUTATION OR GENETIC ENGINEERING; CULTURE MEDIA
- C12N9/00—Enzymes; Proenzymes; Compositions thereof; Processes for preparing, activating, inhibiting, separating or purifying enzymes
-
- C—CHEMISTRY; METALLURGY
- C07—ORGANIC CHEMISTRY
- C07K—PEPTIDES
- C07K2319/00—Fusion polypeptide
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12N—MICROORGANISMS OR ENZYMES; COMPOSITIONS THEREOF; PROPAGATING, PRESERVING, OR MAINTAINING MICROORGANISMS; MUTATION OR GENETIC ENGINEERING; CULTURE MEDIA
- C12N2310/00—Structure or type of the nucleic acid
- C12N2310/30—Chemical structure
- C12N2310/32—Chemical structure of the sugar
- C12N2310/321—2'-O-R Modification
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12N—MICROORGANISMS OR ENZYMES; COMPOSITIONS THEREOF; PROPAGATING, PRESERVING, OR MAINTAINING MICROORGANISMS; MUTATION OR GENETIC ENGINEERING; CULTURE MEDIA
- C12N2310/00—Structure or type of the nucleic acid
- C12N2310/50—Physical structure
- C12N2310/53—Physical structure partially self-complementary or closed
Definitions
- the invention relates to the genetic modification of plants. Particularly, the control of gene integration and expression in plants is provided.
- Genetic modification techniques enable one to insert exogenous nucleotide sequences into an organism's genome.
- a number of methods have been described for the genetic modification of plants. All of these methods are based on introducing a foreign DNA into the plant cell, isolation of those cells containing the foreign DNA integrated into the genome, followed by subsequent regeneration of a whole plant. Unfortunately, such methods produce transformed cells that contain the introduced foreign DNA inserted randomly throughout the genome and often in multiple copies.
- the random insertion of introduced DNA into the genome of host cells can be lethal if the foreign DNA happens to insert into, and thus mutate, a critically important native gene.
- the expression of an inserted foreign gene may be influenced by "position effects" caused by the surrounding genomic DNA.
- the gene is inserted into sites where the position effects are strong enough to prevent the synthesis of an effective amount of product from the introduced gene.
- overproduction of the gene product has deleterious effects on the cell.
- Transgene expression is typically governed by the sequences, including promoters and enhancers, which are physically linked to the transgene.
- transgenes could be activated or inactivated where the sequences that control transgene expression can be altered by either removing sequences present in the original transgene or by inserting additional sequences into the transgene.
- homologous recombination is an essential event participating in processes like DNA repair and chromatid exchange during mitosis and meiosis. Recombination depends on two highly homologous extended sequences and several auxiliary proteins. Strand separation can occur at any point between the regions of homology, although particular sequences may influence efficiency. These processes can be exploited for a targeted integration of transgenes into the genome of certain cell types. Even with the advances in genetic modification of higher plants, the major problems associated with the conventional gene transformation techniques have remained essentially unresolved as to the problems discussed above relating to variable expression levels due to chromosomal position effects and copy number variation of transferred genes.
- compositions and methods for the targeted integration of nucleotide sequences into a transformed plant are provided.
- the compositions comprise transfer cassettes which are flanked by non-identical recombination sites.
- the methods find use in targeting the integration of nucleotide sequences o f interest to a specific chromosomal site, finding optimal integration sites in a plant genome, comparing promoter activity in transformed plants, engineering chromosomal rearrangements, and other genetic manipulation of plants.
- Novel minimal recombination sites FRT are provided for use in the methods of the invention.
- targeting cassettes and transgenic plants and plant cells containing corresponding non-identical recombination sites are also provided.
- Figure 2 provides a construct of the representative plasmid PHP 10616.
- compositions and methods for the directional, targeted integration of exogenous nucleotides into a transformed plant are provided.
- the methods use novel recombination sites in a gene targeting system which facilitates directional targeting of desired genes and nucleotide sequences into corresponding recombination sites previously introduced into the target plant genome.
- a nucleotide sequence flanked by two non-identical recombination sites is introduced into the target organism's genome establishing a target site for insertion of nucleotide sequences of interest.
- a second construct, or nucleotide sequence of interest, flanked by corresponding recombination sites as those flanking the target site is introduced into the stably transformed plant or tissues in the presence of a recombinase protein.
- This process results in exchange of the nucleotide sequences between the non-identical recombination sites of the target site and the transfer cassette.
- the transformed plant may comprise multiple target sites; i.e.
- target site in the transformed plant is intended a DNA sequence that has been inserted into the transformed plant's genome and comprises non-identical recombination sites.
- recombination sites for use in the invention are known in the art and include FRT sites (See, for example, Schlake and Bode (1994) Biochemistry 33: 12746- 12751; Huang et al. (1991) Nucleic Acids Research 19:443-448; Paul D. Sadowski (1995) In Progress in Nucleic Acid Research and Molecular Biology vol. 51, pp. 53-91; Michael M.
- the two-micron plasmid found in most naturally occurring strains of Saccharom ces cerevisiae encodes a site-specific recombinase that promotes an inversion of the DNA between two inverted repeats. This inversion plays a central role in plasmid copy-number amplification.
- the protein, designated FLP protein catalyzes site-specific recombination events.
- the nrinimal recombination site (FRT, SEQ ID NO 1) has been defined and contains two inverted 13 -base pair (bp) repeats surrounding an asymmetric 8- bp spacer.
- the FLP protein cleaves the site at the junctions of the repeats and the spacer and is covalently linked to the DNA via a 3' phosphate.
- Site specific recombinases like FLP cleave and religate DNA at specific target sequences, resulting in a precisely defined recombination between two identical sites.
- the system needs the recombination sites and the recombinase. No auxiliary factors are needed.
- the entire system can be inserted into and function in plant cells.
- the yeast FLP ⁇ FRT site specific recombination system has been shown to function in plants. To date, the system has been utilized for excision of unwanted DNA. See, Lyznik et at. (1993) Nucleic Acid Res. 21:969-975.
- the present invention utilizes non-identical FRTs for the exchange, targeting, arrangement, insertion and control of expression of nucleotide sequences in the plant genome.
- a transformed organism of interest particularly a plant, containing a target site integrated into its genome is needed.
- the target site is characterized by being flanked by non-identical recombination sites.
- a targeting cassette is additionally required containing a nucleotide sequence flanked by corresponding non-identical recombination sites as those sites contained in the target site of the transformed organism.
- a recombinase which recognizes the non-identical recombination sites and catalyzes site-specific recombination is required. It is recognized that the recombinase can be provided by any means known in the art.
- flanking recombination sites are not identical in sequence and will not recombine or recombination between the sites will be minimal. That is, one flanking recombination site may be a FRT site where the second recombination site may be a mutated FRT site.
- non-identical recombination sites used in the methods of the invention prevent or greatly suppress recombination between the two flanking recombination sites and excision of the nucleotide sequence contained therein. Accordingly, it is recognized that any suitable non-identical recombination sites may be utilized in the invention, including FRT and mutant FRT sites, FRT and lox sites, lox and mutant lox sites, as well as other recombination sites known in the art.
- suitable non-identical recombination site implies that in the presence of active recombinase, excision of sequences between two non-identical recombination sites occurs, if at all, with an efficiency considerably lower than the recombinationally-mediated exchange targeting arrangement of nucleotide sequences into the plant genome.
- suitable non-identical sites for use in the invention include those sites where the efficiency of recombination between the sites is low; for example, where the efficiency is less than about 30 to about 50%, preferably less than about 10 to about 30%, more preferably less than about 5 to about 10%.
- the recombination sites in the targeting cassette correspond to those in the target site of the transformed plant. That is, if the target site of the transformed plant contains flanking non-identical recombination sites of FRT and a mutant FRT, the targeting cassette will contain the same FRT and mutant FRT non-identical recombination sites.
- the recombinase which is used in the invention, will depend upon the recombination sites in the target site of the transformed plant and the targeting cassette. That is, if FRT sites are utilized, the FLP recombinase will be needed.
- the Cre recombinase is required. If the non-identical recombination sites comprise both a FRT and a lox site, both the FLP and
- the FLP recombinase is a protein which catalyzes a site-specific reaction that is involved in amplifying the copy number of the two micron plasmid of S. cerevisiae during DNA replication. FLP protein has been cloned and expressed. See, for example, Cox (1993) Proc. Natl. Acad. Sci. U.S.A. 80:4223-4227.
- the FLP recombinase for use in the invention may be that derived from the genus Saccharomyces . It may be preferable to o synthesize the recombinase using plant preferred codons for optimum expression in a plant of interest.
- Cre catalyzes site-specific recombination between two lox sites.
- the Cre recombinase is known in the art. See, for example, Guo et al. (1997) Nature 389:40-46; Abremski et al. (1984) J. Biol. Chem. 259: 1509-1514; Chen et al. (1996) Somat. Cell Mol. Genet. 22:477-488; and Shaikh et al. ⁇ 1911) J. Biol. Chem. 272:5695-5702. All of which are herein incorporated by reference. Such Cre sequence may also be synthesized using plant preferred codons.
- nucleotide sequences to be inserted in the plant genome may be optimized for increased expression in the transformed plant.
- mammalian, yeast, or bacterial genes are used in the invention, they can be synthesized using plant preferred codons for improved expression. It is recognized that for expression in monocots, dicot genes can also be synthesized using monocot preferred codons. Methods are available in the art for synthesizing plant preferred genes. See, for example, U.S. Patent Nos. 5,380,831, 5,436, 391, and Murray et al. (1989) Nucleic Acids Res. 77:477-498, herein incorporated by reference.
- the plant preferred codons may be determined from the codons utilized more frequently in the proteins expressed in the plant of interest. It is recognized that monocot or dicot preferred sequences may be constructed as well as plant preferred sequences for particular plant species. See, for example, EPA 0359472; EPA 0385962; WO 91/16432; Perlak et al. (1991) Proc. Natl. Acad. Sci. USA, 85:3324-3328; and Murray et al. (1989) Nucleic Acids Research, 17: 477-498. U.S. Patent No. 5,380,831; U.S. Patent No. 5,436,391; and the like, herein incorporated by reference. It is further recognized that all or any part of the gene sequence may be optimized or synthetic. That is, fully optimized or partially optimized sequences may also be used.
- Additional sequence modifications are known to enhance gene expression in a cellular host and can be used in the invention. These include elimination of sequences encoding spurious polyadenylation signals, exon-intron splice site signals, transposon-like repeats, and other such well-characterized sequences, which may be deleterious to gene expression.
- the G-C content of the sequence may be adjusted to levels average for a given cellular host, as calculated by reference to known genes expressed in the host cell. When possible, the sequence is modified to avoid predicted hairpin secondary mRNA structures.
- the present invention also encompasses novel FLP recombination target sites (FRT).
- the FRT (SEQ ID NO 1) has been identified as a minimal sequence comprising two 13 base pair repeats, separated by an 8 base spacer, as follows:
- nucleotides within the brackets indicate the spacer region.
- the nucleotides in the spacer region can be replaced with a combination of nucleotides, so long as the two 13- base repeats are separated by eight nucleotides. It appears that the actual nucleotide sequence of the spacer is not critical, however for the practice of the invention, some substitutions of nucleotides in the space region may work better than others.
- the eight base pair spacer is involved in DNA-DNA pairing during strand exchange. The asymmetry of the region determines the direction of site alignment in the recombination event, which will subsequently lead to either inversion or excision. As indicated above, most of the spacer can be mutated without a loss of function. See, for example, Schlake and Bode (1994) Biochemistry 33:12746-12751, herein incorporated by reference.
- Novel FRT mutant sites are provided for use in the practice of the methods of the present invention. Such mutant sites may be constructed by PCR-based mutagenesis. While mutant FRT sites (SEQ ID Nos 2, 3, 4 and 5) are provided herein, it is recognized that other mutant FRT sites may be used in the practice of the invention. The present invention is not the use of a particular FRT or recombination site, but rather that nonidentical recombination sites or FRT sites can be utilized for targeted insertion and expression of nucleotide sequences in a plant genome. Thus, other mutant FRT sites can be constructed and utilized based upon the present disclosure.
- nucleotide sequence of the transfer cassette located between the flanking recombination sites is exchanged with the nucleotide sequence of the target site located between the flanking recombination sites. In this manner, nucleotide sequences of interest may be precisely incorporated into the genome of the host.
- target sites can be constructed having multiple non-identical recombination sites.
- genes or nucleotide sequences can be stacked or ordered at precise locations in the plant genome.
- additional recombination sites may be introduced by incorporating such sites within the nucleotide sequence of the transfer cassette and the transfer of the sites to the target sequence.
- Another variation includes providing a promoter or transcription initiation region operably linked with the target site in an organism.
- the promoter will be 5 ' to the first recombination site.
- advantages of the present system include the ability to reduce the complexity of integration of trans-genes or transferred DNA in an organism by utilizing transfer cassettes as discussed above and selecting organisms with simple integration patterns.
- preferred sites within the genome can be identified by comparing several transformation events.
- a preferred site within the genome includes one that does not disrupt expression of essential sequences and provides for adequate expression of the transgene sequence.
- the methods of the invention also provide for means to combine multiple cassettes at one location within the genome. See, for example, Figure 1. Recombination sites may be added or deleted at target sites within the genome.
- a plant can be stably transformed to harbor the target site in its genome.
- the recombinase may be transiently expressed or provided.
- a nucleotide sequence capable of expressing the recombinase may be stably integrated into the genome of the plant.
- the transfer cassette flanked by corresponding nonidentical recombination sites, is inserted into the transformed plant's genome.
- the components of the system may be brought together by sexually crossing transformed plants.
- a transformed plant, parent one, containing a target site integrated in its genome can be sexually crossed with a second plant, parent two, that has been genetically transformed with a transfer cassette containing flanking non-identical recombination sites, which correspond to those in plant one.
- Either plant one or plant two contains within its genome a nucleotide sequence expressing recombinase.
- the recombinase may be under the control of a constitutive or inducible promoter.
- Inducible promoters include heat-inducible promoters, estradiol-responsive promoters, chemical inducible promoters, and the like.
- Pathogen inducible promoters include those from pathogenesis-related proteins (PR proteins), which are induced following infection by a pathogen; e.g., PR proteins, SAR proteins, beta-l,3-glucanase, chitinase, etc. See, for example, Redolfi et al. (1983) Neth. J. Plant Pathol. 89:245-254; Uknes et al. (1992) The Plant Cell 4: 645-656; and Van Loon (1985) Plant Mol. Virol. 4: 111-116. In this manner, expression of recombinase and subsequent activity at the recombination sites can be controlled.
- PR proteins pathogenesis-related proteins
- Such promoters include, but are not limited to 35S promoter of cauliflower mosaic virus
- ubiquitin promoter (Christensen et al. (1992) Plant Mol. Biol. 18:675-689), promoters from genes such as ribulose bisphosphate carboxy lase (De Almeida et al. (1989) Mol. Gen. Genet. 218:78-98), actin (McElroy et al. (1990) Plant J. 2:163-171), histone, DnaJ (Baszczynski et al. (1997) Maydica 42: 189-201), and the like.
- compositions and methods of the invention find use in targeting the integration of transferred nucleotide sequences to a specific chromosomal site.
- the nucleotide sequence may encode any nucleotide sequence of interest. Particular genes of interest include those which provide a readily analyzable functional feature to the host cell and/or organism, such as marker genes, as well as other genes that alter the phenotype of the recipient cells, and the like. Thus, genes effecting plant growth, height, susceptibility to disease, insects, nutritional value, and the like may be utilized in the invention.
- the nucleotide sequence also may encode an 'antisense' sequence to turn off or modify gene expression.
- nucleotide sequences will be utilized in a functional expression unit or cassette.
- functional expression unit or cassette is intended, the nucleotide sequence of interest with a functional promoter, and in most instances a termination region.
- the nucleic acid of interest is transferred or inserted into the genome as a functional expression unit.
- the nucleotide sequence may be inserted into a site within the genome which is 3' to a promoter region. In this latter instance, the insertion of the coding sequence 3' to the promoter region is such that a functional expression unit is achieved upon integration.
- the nucleic acid encoding target sites and the transfer cassettes, including the nucleotide sequences of interest can be contained within expression cassettes.
- the expression cassette will comprise a transcriptional initiation region, or promoter, operably linked to the nucleic acid encoding the peptide of interest.
- Such an expression cassette is provided with a plurality of restriction sites for insertion of the gene or genes of interest to be under the transcriptional regulation of the regulatory regions.
- the transcriptional initiation region, the promoter may be native or homologous or foreign or heterologous to the host, or could be the natural sequence or a synthetic sequence. By foreign is intended that the transcriptional initiation region is not found in the wild-type host into which the transcriptional initiation region is introduced. Either a native or heterologous promoter may be used with respect to the coding sequence of interest.
- the transcriptional cassette will include in the 5 '-3' direction of transcription, a transcriptional and translational initiation region, a DNA sequence of interest, and a transcriptional and translational termination region functional in plants.
- the termination region may be native with the transcriptional initiation region, may be native with the DNA sequence of interest, or may be derived from another source.
- Convenient termination regions are available from the potato proteinase inhibitor (Pinll) gene or from Ti-plasmid of A. tumefaciens, such as the octopine synthase and nopaline synthase termination regions.
- the expression cassettes may additionally contain 5' leader sequences in the expression cassette construct.
- leader sequences can act to enhance translation.
- Translation leaders are known in the art and include: picornavirus leaders, for example, EMCV leader (Encephalomyocarditis 5' noncoding region) (Elroy-Stein, O., Fuerst, T.R., and Moss, B. (1989) PNAS USA, 86:6126-6130); potyvirus leaders, for example, TEV leader (Tobacco Etch Virus) (Allison et al.
- MDMV leader Mainze Dwarf Mosaic Virus
- Virology 154:9-20
- human immunoglobulin heavy-chain binding protein BiP
- untranslated leader from the coat protein mRNA of alfalfa mosaic virus AMV RNA 4
- TMV tobacco mosaic virus leader
- MCMV chlorotic mottle virus leader
- the expression cassettes may contain one or more than one gene or nucleic acid sequence to be transferred and expressed in the transformed plant. Thus, each nucleic acid sequence will be operably linked to 5' and 3 ' regulatory sequences. Alternatively, multiple expression cassettes may be provided.
- the expression cassette will comprise a selectable marker gene for the selection of transformed cells.
- Selectable marker genes are utilized for the selection of transformed cells or tissues. See generally, G. T. Yarranton (1992) Curr. Opin. Biotech., 5:506-511;
- the methods of the invention can also be utilized to find optimal integration sites within a plant genome.
- a plant is transformed with an expression cassette comprising a selectable marker gene.
- the expression cassette is a target site as the marker gene is flanked by non-identical recombination sites.
- Transformed protoplast, tissues, or whole plants can be tested to determine the levels of activity of the inserted gene.
- preferred integration sites may be found wherein the gene is expressed at high or acceptable levels.
- These plants can then be utilized with subsequent retargeting techmques to replace the marker gene with other genes or nucleotide sequences of interest.
- multiple genes may be inserted at the optimal site for expression. See, for example, Figure 2 which sets forth one scheme for gene stacking utilizing site-specific integration using the FRT/FLP system.
- compositions of the present invention including, for example, comparing promoter activity in a transformed plant.
- promoter activity could not accurately be assessed and compared because the chimeric genes were inserted at different locations within the plant genome. Such chromosomal locations affected activity.
- a direct comparison of promotor activity in a defined chromosomal context is possible.
- enhanced activity of genes can be achieved by selecting optimal chromosomal sites as well as optimal promoters for expression in the plant cell.
- the present invention may be used for transformation of any plant species, including but not limited to corn ⁇ Zea mays), canola ⁇ Brassica napus, Brassica rapa ssp.), alfalfa ⁇ Medicago sativa), rice ⁇ Oryza sativa), rye ⁇ Secale cereale), sorghum ⁇ Sorghum bicolor, Sorghum vulgare), sunflower ⁇ Helianthus annuus), wheat ⁇ Triticum aestivum), soybean ⁇ Glycine max), tobacco ⁇ Nicotiana tabacum), potato ⁇ Solanum tuberosum), peanuts ⁇ Arachis hypogaea), cotton ⁇ Gossypium hirsutum), sweet potato ⁇ Ipomoea batatus), cassava ⁇ Manihot esculenta), coffee ⁇ Cofea spp.), coconut ⁇ Cocos nucifera), pineapple ⁇ Ananas comosus), citrus trees ⁇ Cit
- Vegetables include tomatoes (Jy coper sicon esculentum), lettuce (e.g., Lactuca sativa), green beans ⁇ Phaseolus vulgaris), lima beans ⁇ Phaseolus limensis), peas ⁇ Lathy rus spp.) and members of the genus Cucumis such as cucumber (C. sativus), cantaloupe (C. cantalupensis), and musk melon (C. me/ ⁇ ).
- Ornamentals include azalea ⁇ Rhododendron spp.), hydrangea ⁇ Macrophylla hydrangea), hibiscus ⁇ Hibiscus rosasanensis), roses (R ⁇ r ⁇ spp.), tulips ⁇ Tulipa spp.), daffodils ⁇ Narcissus spp.), petunias ⁇ Petunia hybrida), carnation ⁇ Dianthus caryophyllus) , poinsettia ⁇ Euphorbia pulcherrima), and chrysanthemum.
- Conifers which may be employed in practicing the present invention include, for example, pines such as loblolly pine ⁇ Pinus taeda), slash pine ⁇ Pinus elliotii), ponderosa pine ⁇ Pinus ponderosa), lodgepole pine ⁇ Pinus contorta), and Monterey pine ⁇ Pinus radiata); Douglas-fir ⁇ Pseudotsuga menziesii); Western hemlock ⁇ Tsuga canadensis); Sitka spruce ⁇ Picea glauca); redwood ⁇ Sequoia sempervirens); true firs such as silver fir ⁇ Abies amabilis) and balsam fir ⁇ Abies balsamea); and cedars such as Western red cedar ⁇ Thuja plicata) and Alaska yellow-cedar ⁇ Chamaecyparis nootkatensis).
- pines such as loblolly pine ⁇ Pin
- plants of the present invention are crop plants (for example, corn, alfalfa, sunflower, canola, soybean, cotton, peanut, sorghum, wheat, tobacco, etc.), more preferably corn and soybean plants, yet more preferably corn plants. It is recognized that the methods of the invention may be applied in any plant system. Methods for transformation of plants are known in the art.
- Transformation protocols may vary depending on the type of plant or plant cell, i.e. , monocot or dicot, targeted for transformation. Suitable methods of transfomiing plant cells include microinjection (Crossway et al. (1986) Biotechniques 4:320-334), electroporation (Riggs et al. (1986) Proc. Natl. Acad. Sci. USA, 85:5602-5606, Agrobacterium mediated transformation (Hinchee et al. (1988) Biotechnology, 6:915-921), direct gene transfer (Paszkowski et al.
- the cells which have been transformed may be grown into plants in accordance with conventional approaches. See, for example, McCormick et al. (1986) Plant Cell Reports, 5:81-84. These regenerated plants may then be pollinated with either the same transformed strain or different strains, and the resulting hybrid having the desired phenotypic characteristic identified. Two or more generations may be grown to ensure that the subject phenotypic characteristic is stably maintained and inherited and then seeds harvested to ensure the desired phenotype or other property has been achieved.
- Agrobacterium-mediated transformation may be preferred.
- Agrobacterium-mediated transformation generally tends to insert a lower copy number of transferred DNA than does particle bombardment or other transformation means.
- the general present invention provides a procedure for using existing and novel FRT sites in a new gene targeting system which facilitates directional retargeting of desired genes into FRT sites previously introduced in the target organism's genome.
- the novel FRT sites differ from previously described FRT sites in the sequence of the 8 bp spacer regions of the FRT sites.
- Previous publications also have shown that in the presence of FLP protein, recombination of sequences between two FRT sites occurs efficiently only with two identical FRT sites. See for example Umlauf and Cox (1988) Embo J. 7: 1845- 1852; Schlake and Bode (1994) Biochem. 33: 12746-12751.
- a gene or DNA sequence is flanked by two non-identical FRT sites and introduced into a target organism's genome.
- the enclosed gene can be a selectable marker, thereby allowing selection for successfully introduced sequences.
- Molecular characterization confirms integration of desired sequences including complete FRT sites. Listed below are generic examples of vector constructions useful in practicing the invention:
- FRTa and FRTb are two examples of non-identical FRT sites.
- PI, P2 and P3 are different promoters, Gl, G2, and G3 are different genes, TI, T2 and T3 are different terminators.
- ATG is the start of translation codon for the subsequent gene.
- the designation noATG indicates that particular gene is devoid of the ATG translation start codon.
- a to F are preferred configurations for testing new FRT sites for ability to recombine sequences between them; the desired situation being that when two of the same site are used, recombination is efficient and that when two different sites are used, no recombination between them takes place in the presence of FLP protein.
- G to J are preferred configurations for general use in developing lines for retargeting. It is understood that any number of genes or other combinations of sequences can be assembled for use as part of this invention.
- K to N are possible configurations that could be used also.
- a second construct flanked by the same FRT sites used to flank the sequences in the first construct above is introduced into the stably transformed tissues in conjunction with the expression of FLP protein.
- the new vector constructs can be, but are not limited to the following:
- the FLP protein can be supplied by a) co-transforming with a plasmid carrying a gene encoding FLP; b) co-introducing FLP mRNA or protein directly; c) using a line for the initial transformation that expresses FLP either constitutively or following induction; or d) growing out the plants carrying the initial targeted vectors, crossing to plants that express active FLP protein and selecting events in the progeny.
- sequence O above is introduced into a line containing a copy of sequence G stably integrated in the genome, in the presence of functional FLP protein. Recombination takes place between identical FRT sites such that the sequence between FRT sites in O replaces the sequence between the corresponding FRT sites of sequence G, thereby yielding a directionally targeted reintegrated new sequence.
- the new gene in O is now driven off of the PI promoter in G.
- the purpose for designing some of the constructs without an ATG start codon on the gene is so that if random integration occurs, there is an extremely low probability of expression of the introduced gene, since in order for this to happen, the fragment would need to integrate behind an endogenous promoter region and in the correct reading frame.
- DNA fragments containing novel FRT sequences were constructed either by synthesizing, annealing and ligating complementary oligonucleotides or by creating primers for PCR amplification (Mullis and Faloona, 1987) of a DNA product containing the new FRT sequence near the 5' end of the PCR product.
- the newly constructed FRT product includes flanking restriction sites useful for cloning into plant expression units. In general, the 5' end is flanked by an Nhel site and a terminal Ncol site.
- the Ncol site includes the bases ATG, which are advantageously used in newly developed vector constructs as the recognition sequence to initiate an open reading frame.
- the Nhel site is used for cloning thereby eliminating the upstream ATG in the process.
- a restriction site is included at the 3' end of the FRT sequence.
- the wild type FRT site (designated FRT1 here) is cloned with a flanking Bglll site
- the FRT5 site spacer TTCAAAAG
- the FRT6 site (spacer TTCAAAAA) has an Aatll site
- the FRT7 site spacer TTCAATAA) has an Spel site.
- the outermost flanking restriction site is an Xhol site and is used to clone a gene of interest into the open reading frame.
- FRT sites as designed and/or used in the present invention example are depicted below with positions of restriction sites, repeats and spacer regions indicated.
- FRT5 (SEQ ID NO 3)
- FRT6 (SEQ ID NO 4)
- FRT7 (SEQ ID NO 5)
- PCR or standard mutagenesis protocols were used to create an Xhol site overlapping the start of a gene sequence to be used for cloning downstream of the FRT site, thereby converting the ATG start codon to GTG.
- Ligation of an FRT to the mutated gene sequence at Xhol creates a new open reading frame initiating 5' to the FRT.
- a second FRT sequence can be cloned downstream of the terminator using a variety of methods including PCR or ligation.
- the FRT/gene/terminator/FRT unit can then be used to make target or substrate constructs. Targets are created by inserting a promoter at the Ncol site upstream of the first
- FRT This maintains a complete open reading frame of the FRT/gene fusion.
- target constructs are for use in transformation experiments to create desirable 'target lines'.
- Substrate vectors are constructed by cloning with the Nhel site to truncate the start codon of the FRT /gene unit, thereby eliminating the proper open reading frame.
- These substrate vectors are used in experiments designed to retarget a new gene flanked by FRT sites into the corresponding FRT sites previously introduced in the target lines. In either case, to create multiple gene cassettes, additional promoter/gene/terminator units are inserted between the terminator and the second FRT in either target or substrate molecules.
- Example 3 Demonstration of functionality of novel FRT sites and requirement for two identical sites for efficient recombination of DNA sequences positioned between two FRT sites.
- Plasmids containing two identical or two different FRT sequences were assayed for efficiency of recombination of sequences between the FRT sites by transformation into 294-FLP, a version of the E. coli strain MM294 with FLP recombinase integrated into the lacZ locus (Buchholz et al. 1996). Strains were grown overnight at 37°C with shaking, allowing for constitutive expression of FLP recombinase in the cultures. The plasmid DNA was isolated using standard procedures and digested with restriction enzymes that create novel restriction fragments following FLP mediated recombination. The extent of recombination between FRT sites was estimated by examining banding patterns on an agarose gel. Table 1 summarizes data from the gel analysis. Table 1
- Example 4 Introduction of DNA sequences which include novel non-identical FRT sites into plant cells, generation and recovery of stable transgenic events ('target lines'), preservation of 'target lines' and regeneration of plants.
- a number of stable transgenic events carrying FRT target sites were produced. These target lines were generated by introducing one of a series of constructs including, for example, PHP9643, PHP10616, PHP11407, PHP11410, PHP11457, PHP11599, PHP11893 or PHP14220 (See Table 2) into corn cells, either by particle bombardment, as described in Register et al. (1994) Plant Mol. Biol. 25:951-961 or via Agrobacterium co- cultivation as described by Heath et al. (1997) Mol. Plant-Microbe Interact. 10:22-221; Hiei et al. (1994) Plant J. 6:271-282 and Ishida et al. (1996) Nat. Biotech. 74:745-750, and in U.S.
- PHP 10616 has the mo-PAT (U.S. Provisional Patent Application Serial No. 60/035,560 to "Methods for Improving Transformation Efficiency", filed January 14, 1997) coding sequence fused in frame at the Xhol site flanking FRTl (see above, SEQ ID NO 2).
- PHP11407 and PHP11893 have GFPm-C3 (PCT/US97/07688 filed May 1, 1997 from Provisional Application 60/016,345 filed May 1, 1996) containing the second intron from potato ST-LSl (Vancanneyt et al. (1990) Mol. Gen. Genet.
- the potato proteinase inhibitor II (Pinll) terminator (bases 2 to 310 from An et al. (1989) Plant Cell 1: 115-122) was ligated downstream of the coding sequences.
- PHP10616 has an FRT5 sequence (SEQ ID NO 3) cloned downstream of the Pinll terminator.
- the second expression units have the maize ubiquitin promoter or alternatively either the enhanced or the standard versions of the cauliflower mosaic virus 35S promoter.
- the standard 35S promoter includes bases -421 to +2 (from Gardner et al. (1981) Nucl. Acids Res. 9:2871-2888), and the enhanced version has a duplication of bases -421 to -90 upstream of this standard 35S promoter.
- the 79 bp tobacco mosaic virus leader O' (Gallie et al. (1987) Nucl. Acids Res. 15:3257-3273) is inserted downstream of the 35S promoter followed by the first intron of the maize alcohol dehydrogenase ADH1-S gene (Dennis et al. (1984) Nucl.
- Coding sequences in these second expression units include either mo-PAT, bar (Thompson et al. (1987) EMBO J. 6:2519-2523), or HM7 (Johal and Briggs, Science 258:985-987) genes followed by either the Pinll terminator or the 35S terminator (nucleotides 7487-7639 in Gardner et al. (1981) Nucl. Acids Res. 9:2871-2888). Varying FRT sites are ligated downstream of the terminators as shown in the table.
- a third expression unit is present in P ⁇ P9643 and has an FRTl /GFPm fusion cloned using the flanking Nhel site of FRTl (SEQ ID NO 2) to remove the ATG start codon of GFPm, thereby making it non-functional in the existing construct, but where correct excision of sequences between FRTl (SEQ ID NO 2) sites can bring the GFPm in frame with the ubiquitin promoter and ATG of the first expression unit, thereby making it functional.
- Downstream of GFPm is the Pinll terminator followed by an FRT5 sequence (SEQ ID NO 3).
- PHP9643 was cloned into a pUC derived plasmid backbone. All other vectors were cloned into a pSBl l (See, for example, EPA0672752A1, EPA0604662A1, EPA0687730A1 and U.S. Patent No. 5,591,616) type plasmid with the expression units contained between the TDNA border sequences. All are oriented with expression unit one adjacent to the right border.
- the pSB 11 -based plasmids were integrated into the super binary plasmid pSBl (See, for example, EPA0672752A1, EPA0604662A1, EPA0687730A1 and U.S. Patent No.
- E. coli strain HB101 containing the pSBll derivatives was mated with Agrobacterium strain LBA4404 harboring pSBl to create the cointegrate plasmids PHP10616, PHP11407, PHP11410, PHP11457, PHP11599, PHP11893 and PHP14220 in Agrobacterium (by the method of Ditta et al. (1980) Proc. Natl. Acad. Sci. USA 77:7347-7351).
- the cointegrates were verified by Agrobacterium resistance to spectinomycin and Sail restriction digests.
- Table 2 also includes one example of a vector for creating a target line where the FRT sites are inserted in the maize ubiquitin intron (last entry) as an alternative location for placement of FRT or other target sites.
- the extent of intra-plasmid recombination was examined in plants using the FRT excision constructs described in Table 3 below.
- the vectors PHP 10968, PHP 10998, PHP10969, PHP11272, PHP11243, PHP11244, PHP12140, PHP12141, PHP12156, and PHP12157 were constructed by ligating the maize Ubiquitin promoter upstream of FRT sequences using Ncol or other sites that maintained the ATG start codon.
- the FRT sequence was fused in frame at the flanking Xhol site to a GFPm sequence containing a serine to threonine mutation at amino acid residue 65 in the wild type sequence (new sequence termed GFPm-S65T).
- the pinll terminator was cloned downstream of GFPm.
- the second expression unit consists of a promoterless FRT, cloned with the 5' flanking
- the vector backbone is a pUC derived plasmid in all cases.
- PHP5096 the FLPm expression vector that was used in experiments with the excision and substrate vectors, consists of the maize Ubiquitin promoter cloned upstream of the FLPm coding sequence (U.S. Patent Application Serial No. 08/972,258 to "Novel Nucleic Acid Sequence
- FRT substrate vectors are constructed as promoterless FRT/gene fusions cloned using the 5' flanking Nhel site of the FRT to remove the ATG start codon.
- Genes fused in frame to the FRT with the flanking Xhol site include one of several scorable or selectable marker genes such as aadA (Svab et al. (1990) Plant Mol. Biol. 14: 197-205), uidA, GFPm, GFPm-C3/intron or bar and are followed by a pinll terminator.
- plasmids contain a single expression unit and the second heterologous FRT site is cloned downstream of the pinll terminator.
- Substrate plasmids PHP10859, PHP10997, PHPl 1204, PHPl 1699, and PHP12190 have in addition to the first expression unit described above, a second unit consisting of the maize ubiquitin promoter, the enhanced 35S promoter or a chimeric promoter consisting of the 35S enhancer region cloned upstream of a synthetic core promoter termed Rsyn7 (U.S. Patent Application Serial No.
- PHPl 1003 and PHPl 1809 contain three expression units.
- the first unit is a promoterless noATG/FRT/gene fusion as described above
- the second unit contains either the chimeric 35S enhancer/Rsyn7 promoter described above or the ZmdJl promoter (Baszczynski et al. (1997) Maydica 42: 189-201) cloned upstream of the GUS coding sequence and the pinll terminator.
- the third expression unit consists of the maize ubiquitin promoter cloned upstream of the HMl coding sequence, pinll terminator and a heterologous FRT sequence. All FRT substrate vectors are cloned into a pUC derived plasmid backbone. Details of the components of these vectors are described in Table 6. Also listed in Table 6 are two vectors with alternative placement of FRT sites in the ubiquitin 5' UTR or intron.
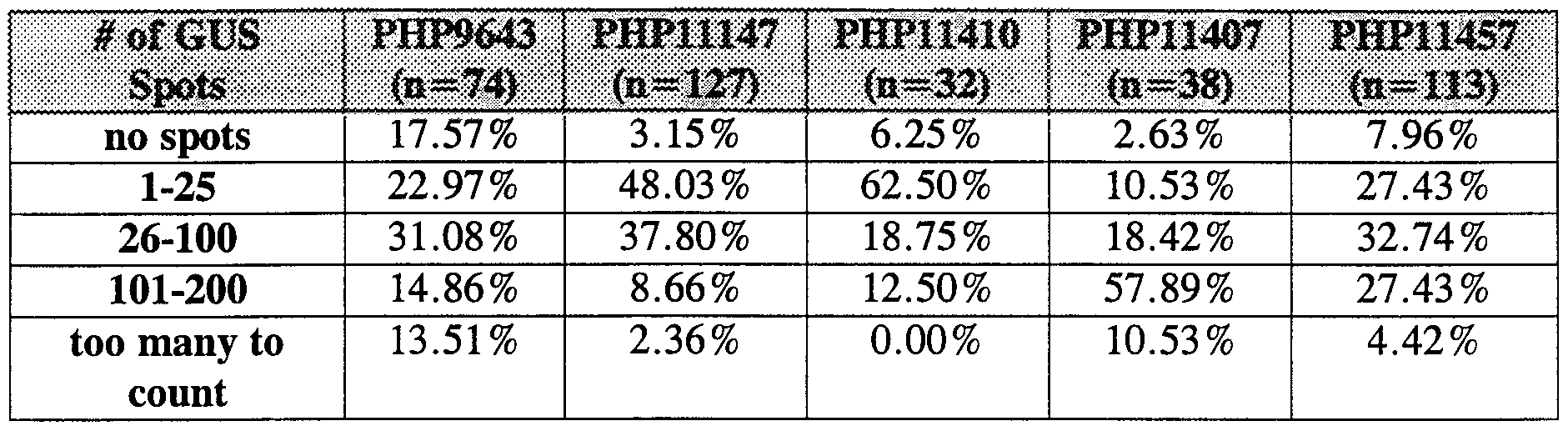
- Results in Table 5 indicate that the frequency and level of GUS expression varies among different events, as might be predicted for genes inserted in different positions in the genome. The prediction is that once a high frequency, high expressing line is identified, that the expression of genes subsequently introduced into those same sites will also be higher than in other lower expressing events.
- Table 8 presents data from stable transgenic events which demonstrate successful and reproducible targeting of introduced sequences to previously created genomic target sites.
- the data shown are for 18 independent target lines, each retargeted with a promoterless GUS construct. Since the bar gene was concurrently introduced on the same plasmid, the proportion of GUS expressing events from the total events recovered on bialophos selection provides a measure of retargeting frequency relative to random integration.
- Example 6 Evaluation of impact of introduced FRT sequences on plant development, gene expression and agronomic performance.
- Example 7 Conversion of an introduced functional FRT sequence into a second non-identical functional FRT sequence
- the target vectors for evaluating this FRT site modification strategy are shown generically below, where PI and P2 represent two different promoters, Gl and G2 represent two genes, and TI and T2 represent two terminator regions; these regions are shown as white boxes. Different FRT sites are indicated and shown as dark boxes.
- One version of the construct incorporates a third unique FRT site downstream of the second gene and is used to evaluate whether the targeted conversion, in this case, of FRT5 to FRT6 (SEQ ID NO 4), also results in conversion of the downstream FRTl (SEQ ID NO
- PI is the maize ubiquitin promoter
- P2 is the enhanced CaMV 35S promoter
- Gl is the uidA (GUS) gene
- G2 is the bar gene
- TI and T2 are pinll terminators. It is understood that based on the various descriptions of vector constructs earlier in this application, a variety of different promoters, genes, terminators or DNA sequences or FRT sites could be used in practicing this component method.
- the DNA cassettes as shown below could be assembled into either a pUC-based plasmid for direct DNA delivery methods (such as particle bombardment) or into a binary vector for Agrobacterium-based transformation as described previously.
- chimeric molecules that would be used to modify a single nucleotide so as to convert the FRT5 (SEQ ID NO 3) site to an FRT6 (SEQ ID NO 4) site in constructs as described above. Both the linear sequence of these chimeric molecules as well as the predicted active form of the molecule (based on the Yoon et al. and Cole-Strauss et al. publications above) are shown. DNA residues are represented in upper case, RNA residues in lower case, and the site to be modified (a single nucleotide difference between FRT5, SEQ ID NO 3, and FRT6, SEQ ID NO 4) is underlined and in bold. Two examples of chimeras are presented below differing in the number of residues downstream of the FRT5 (SEQ ID NO 4) site that would be included in the chimeric molecule design and which would thus determine the specificity to the target sequence.
- Chimeric oligonucleotide linear sequence (sequence includes six target- specific residues downstream of the FRT site being modified in the target construct and should convert only this single specific FRT5, SEQ ID NO 3, site to an FRT6, SEQ ID NO 4, site) 5 ' -
- DNA from those samples that exhibit GUS expression is isolated, amplified by PCR if necessary, and sequenced by standard methods through the region corresponding to the predicted nucleotide conversion. A sufficient stretch of DNA is sequenced to cover the entire originally introduced region of DNA so as to confirm correct and specific conversion.
- Southern analysis and/or sequencing of GUS expressing and non-expressing samples establishes the presence or absence of specific DNA fragments prior to and following chimeric molecule and FLP recombinase delivery, and thus substantiates the visual and biochemical observations made above.
- Described in Figure 1 is one potential strategy for combining or stacking multiple desired transgenes at one genomic location using the non-identical FRT-based system of the present invention. While stacking of genes can be achieved without the use of the targeted FRT conversion method described in this example 7, this latter method extends the capabilities of the system by allowing in vivo conversion of FRT sites to create new sites, rather than re-introducing new FRT sites by transformation.
- an FRT site with an asterisk beside it indicates that it was initially created to be non-functional with respect to recombination between it and the equivalent FRT site without an asterisk, but which upon conversion with the chimeraplasty-based approach described herein renders it capable of recombination with its equivalent non-asterisk counterpart.
- this method also provides a mechanism to recycle selectable markers, as is possible in using the FRT system of the present invention alone.
- a major problem associated with developing the FLP/FRT system for integrating genes into animals or plants stems from the fact that the recombination reaction catalyzed by yeast FLP recombinase is a reversible process (Sadowski (1995) in Progress in Nucleic Acid Research and Molecular Biology 51:53-91).
- recombination should lead to insertion of the new DNA sequences at the endogenous FRT site.
- the reverse reaction would lead to re-excision of the introduced sequences because of recombination between the identical FRT sites.
- the present invention describes the development of a useful new gene targeting system for plants which utilizes the yeast FLP recombinase or a modified FLP recombinase designed to work more efficiently in certain plant species and novel non-identical FRT sites which can be used for directional non-reversible DNA integration. Additionally, described herein is a novel use of accessory technologies such as 'chimeraplasty' permitting in vivo or in vitro modification of DNA sequences, such as FRT sites to further extend the utility of the system. Data provided demonstrate the successful stable integration of DNA sequences between two previously introduced non-identical FRT sites in maize. We show also that the DNA sequences between the FRT sites can be subsequently replaced by a second DNA sequence flanked by the same FRT sites as the first.
- the present invention provides a means for facilitating the introduction of novel genes or DNA sequences into genomic locations previously dete ⁇ nined to be particularly beneficial for gene integration from the perspective of providing suitable levels of stable expression of the introduced gene(s) and not exhibiting deleterious impacts on agronomic characteristics including yield.
- the invention provides a system whereby integration of two or more genes can be targeted to the same genomic location, providing a mechanism for 'gene stacking'.
- this invention also provides an improved method for introducing, maintaining and breeding multiple genetic traits of interest, including agronomic traits, commercially important genes or other heterologous gene products.
- the invention further proposes to use the non-recombination feature of nonidentical FRT sites to allow creation of a set of 'parental' lines, which are initially well- characterized for all the desired expression and performance parameters described above. These lines then serve as the basis for introduction of new traits into the same predefined sites in the genome where the initial genes were introduced. Many fewer events would need to be generated, since integration would preferentially occur in sites shown to express well and have minimal negative impact on performance.
Abstract
Description







Claims
Priority Applications (7)
| Application Number | Priority Date | Filing Date | Title |
|---|---|---|---|
| AT98958629T ATE302271T1 (en) | 1997-11-18 | 1998-11-17 | COMPOSITIONS AND METHODS FOR GENETIC MODIFICATION OF PLANTS |
| AU14629/99A AU760113C (en) | 1997-11-18 | 1998-11-17 | Compositions and methods for genetic modification of plants |
| DE69831265T DE69831265T2 (en) | 1997-11-18 | 1998-11-17 | COMPOSITIONS AND METHODS FOR THE GENETIC MODIFICATION OF PLANTS |
| NZ503859A NZ503859A (en) | 1997-11-18 | 1998-11-17 | Non-identical minimal recombination sites (FRT) which are non-identical for targeted integration of nucleotide sequences into transformed plants |
| EP98958629A EP1034262B1 (en) | 1997-11-18 | 1998-11-17 | Compositions and methods for genetic modification of plants |
| CA002306184A CA2306184C (en) | 1997-11-18 | 1998-11-17 | Compositions and methods for genetic modification of plants |
| AU2003202440A AU2003202440B8 (en) | 1997-11-18 | 2003-03-24 | Compositions and Methods for Genetic Modification of Plants |
Applications Claiming Priority (4)
| Application Number | Priority Date | Filing Date | Title |
|---|---|---|---|
| US6561397P | 1997-11-18 | 1997-11-18 | |
| US6562797P | 1997-11-18 | 1997-11-18 | |
| US60/065,627 | 1997-11-18 | ||
| US60/065,613 | 1997-11-18 |
Publications (1)
| Publication Number | Publication Date |
|---|---|
| WO1999025821A1 true WO1999025821A1 (en) | 1999-05-27 |
Family
ID=26745783
Family Applications (2)
| Application Number | Title | Priority Date | Filing Date |
|---|---|---|---|
| PCT/US1998/024609 WO1999025854A1 (en) | 1997-11-18 | 1998-11-17 | A method for directional stable transformation of eukaryotic cells |
| PCT/US1998/024610 WO1999025821A1 (en) | 1997-11-18 | 1998-11-17 | Compositions and methods for genetic modification of plants |
Family Applications Before (1)
| Application Number | Title | Priority Date | Filing Date |
|---|---|---|---|
| PCT/US1998/024609 WO1999025854A1 (en) | 1997-11-18 | 1998-11-17 | A method for directional stable transformation of eukaryotic cells |
Country Status (11)
| Country | Link |
|---|---|
| US (13) | US6187994B1 (en) |
| EP (3) | EP1034262B1 (en) |
| AT (2) | ATE401410T1 (en) |
| AU (3) | AU760113C (en) |
| CA (2) | CA2305866A1 (en) |
| DE (2) | DE69831265T2 (en) |
| DK (1) | DK1034262T3 (en) |
| ES (2) | ES2245487T3 (en) |
| NZ (2) | NZ503859A (en) |
| PT (1) | PT1034262E (en) |
| WO (2) | WO1999025854A1 (en) |
Cited By (248)
| Publication number | Priority date | Publication date | Assignee | Title |
|---|---|---|---|---|
| WO1999055851A2 (en) * | 1998-04-28 | 1999-11-04 | Novartis Ag | Site-directed transformation of plants |
| WO2001085969A2 (en) * | 2000-05-08 | 2001-11-15 | Pioneer Hi-Bred International, Inc. | Genetic transformation in plants using site-specific recombination and wide hybridization |
| WO2002006500A2 (en) * | 2000-07-18 | 2002-01-24 | Pioneer Hi-Bred International, Inc. | Methods of transforming plants and identifying parental origin of a chromosome in those plants |
| WO2002008409A2 (en) * | 2000-07-21 | 2002-01-31 | The United States Of America, As Represented By The Secretary Of Agriculture | Methods for the replacement, translocation and stacking of dna in eukaryotic genomes |
| WO2003087313A2 (en) | 2002-04-08 | 2003-10-23 | Pioneer Hi-Bred International, Inc. | Enhanced silk exsertion under stress |
| WO2002077246A3 (en) * | 2001-03-23 | 2003-11-20 | Icon Genetics Ag | Site-targeted transformation using amplification vectors |
| US6746870B1 (en) | 1999-07-23 | 2004-06-08 | The Regents Of The University Of California | DNA recombination in eukaryotic cells by the bacteriophage PHIC31 recombination system |
| EP1499720A2 (en) * | 2002-05-03 | 2005-01-26 | Pioneer Hi-Bred International, Inc. | Gene targeting using replicating dna molecules |
| WO2005016504A2 (en) | 2003-06-23 | 2005-02-24 | Pioneer Hi-Bred International, Inc. | Disruption of acc synthase genes to delay senescence in plants |
| WO2005035769A2 (en) | 2003-10-09 | 2005-04-21 | E. I. Du Pont De Nemours And Company | Gene silencing by using micro-rna molecules |
| WO2005059148A1 (en) * | 2003-12-17 | 2005-06-30 | Pioneer Hi-Bred International, Inc. | Recombinase mediated gene traps |
| WO2007011733A2 (en) | 2005-07-18 | 2007-01-25 | Pioneer Hi-Bred International, Inc. | Modified frt recombination sites and methods of use |
| US7193131B2 (en) | 2001-01-19 | 2007-03-20 | Icon Genetics Ag | Processes and vectors for plastid transformation of higher plants |
| WO2007059077A2 (en) | 2005-11-14 | 2007-05-24 | E.I.Du Pont De Nemours And Company | Compositions and methods for altering alpha- and beta-tocotrienol content |
| WO2007079161A2 (en) | 2005-12-30 | 2007-07-12 | Pioneer Hi-Bred International, Inc. | Udp-xylose synthases (uxs) polynucleotides, polypeptides, and uses thereof |
| WO2007103738A2 (en) | 2006-03-01 | 2007-09-13 | Pioneer Hi-Bred International, Inc. | Compositions related to the quantitative trait locus 6 (qtl6) in maize and methods of use |
| WO2008002872A2 (en) | 2006-06-28 | 2008-01-03 | Pioneer Hi-Bred International, Inc. | Soybean event 3560.4.3.5 and compositions and methods for the identification and/or detection thereof |
| US7351877B2 (en) | 2002-03-29 | 2008-04-01 | Syngenta Participations Ag | Lambda integrase mediated recombination in plants |
| US7371923B2 (en) | 2001-07-06 | 2008-05-13 | Icon Genetics Ag | Process of generating transplastomic plants or plant cells devoid of a selection marker |
| EP1961293A2 (en) | 2001-09-27 | 2008-08-27 | Pioneer Hi-Bred International, Inc. | Phytate polynucleotides and methods of use |
| WO2008112970A2 (en) | 2007-03-14 | 2008-09-18 | Pioneer Hi-Bred International, Inc. | Dominant gene suppression transgenes and methods of using same |
| WO2008145629A2 (en) | 2007-05-25 | 2008-12-04 | Cropdesign N.V. | Yield enhancement in plants by modulation of maize alfins |
| EP2003205A2 (en) | 2004-12-28 | 2008-12-17 | Pioneer Hi-Bred International, Inc. | Improved grain quality through altered expression of seed proteins |
| EP2039769A2 (en) | 2000-09-13 | 2009-03-25 | Pioneer Hi-Bred International, Inc. | Antimicrobial peptides and methods of use |
| US7521240B2 (en) | 2001-05-30 | 2009-04-21 | Smithkline Beecham Corporation | Chromosome-based platforms |
| WO2009067580A2 (en) | 2007-11-20 | 2009-05-28 | Pioneer Hi-Bred International, Inc. | Maize ethylene signaling genes and modulation of same for improved stress tolerance in plants |
| EP2112223A2 (en) | 2005-11-10 | 2009-10-28 | Pioneer Hi-Bred International Inc. | DOF (DNA binding with one finger) sequences and method of use |
| EP2113512A2 (en) | 2004-07-02 | 2009-11-04 | Pioneer Hi-Bred International Inc. | Antifungal polypeptides |
| US7652194B2 (en) | 2000-12-08 | 2010-01-26 | Icon Genetics Gmbh | Processes and vectors for producing transgenic plants |
| US7667091B2 (en) | 2001-03-29 | 2010-02-23 | Icon Genetics Gmbh | Method of encoding information in nucleic acids of a genetically engineered organism |
| US7667092B2 (en) | 2001-04-30 | 2010-02-23 | Icon Genetics Gmbh | Processes and vectors for amplification or expression of nucleic acid sequences of interest in plants |
| EP2161340A2 (en) | 1999-12-16 | 2010-03-10 | CropDesign N.V. | Optimized T-DNA transfer and vectors therefor |
| WO2010065867A1 (en) | 2008-12-04 | 2010-06-10 | Pioneer Hi-Bred International, Inc. | Methods and compositions for enhanced yield by targeted expression of knotted1 |
| WO2010078445A1 (en) | 2008-12-31 | 2010-07-08 | Pioneer Hi-Bred International, Inc. | Auxotrophic agrobacterium for plant transformation and methods thereof |
| WO2010080430A1 (en) * | 2008-12-19 | 2010-07-15 | E. I. Du Pont De Nemours And Company | Site-specific integration and stacking of transgenes in soybean via dna recombinase mediated cassette exchange |
| WO2010083328A2 (en) | 2009-01-14 | 2010-07-22 | The Salk Institute For Biological Studies | Methods for screening and compounds that protect against amyloid diseases |
| US7763458B2 (en) | 2000-10-06 | 2010-07-27 | Icon Genetics Gmbh | Vector system for plants |
| WO2010096613A1 (en) | 2009-02-19 | 2010-08-26 | Pioneer Hi-Bred International, Inc. | Blended refuge deployment via manipulation during hybrid seed production |
| WO2010101818A1 (en) | 2009-03-02 | 2010-09-10 | Pioneer Hi-Bred International, Inc. | Nac transcriptional activators involved in abiotic stress tolerance |
| WO2010120862A1 (en) | 2009-04-14 | 2010-10-21 | Pioneer Hi-Bred International, Inc. | Modulation of acc synthase improves plant yield under low nitrogen conditions |
| EP2251349A1 (en) | 2006-04-19 | 2010-11-17 | Pioneer Hi-Bred International, Inc. | Isolated polynucleotide molecules corresponding to mutant and wild-type alleles of the maize D9 gene and methods of use |
| EP2261361A2 (en) | 2005-05-25 | 2010-12-15 | Pioneer Hi-Bred International Inc. | Methods for improving crop plant architecture and yield |
| WO2010147825A1 (en) | 2009-06-09 | 2010-12-23 | Pioneer Hi-Bred International, Inc. | Early endosperm promoter and methods of use |
| EP2275536A1 (en) | 2002-08-06 | 2011-01-19 | Verdia, Inc. | AP1 amine oxidase variants |
| WO2011011273A1 (en) | 2009-07-24 | 2011-01-27 | Pioneer Hi-Bred International, Inc. | The use of dimerization domain component stacks to modulate plant architecture |
| WO2011017492A2 (en) | 2009-08-05 | 2011-02-10 | Pioneer Hi-Bred International, Inc. | Novel eto1 genes and use of same for reduced ethylene and improved stress tolerance in plants |
| WO2011021171A1 (en) | 2009-08-21 | 2011-02-24 | Beeologics, Llc | Preventing and curing beneficial insect diseases via plant transcribed molecules |
| WO2011022597A1 (en) | 2009-08-20 | 2011-02-24 | Pioneer Hi-Bred International, Inc. | Functional expression of shuffled yeast nitrate transporter (ynti) in maize to improve nitrate uptake under low nitrate environment |
| WO2011022608A2 (en) | 2009-08-20 | 2011-02-24 | Pioneer Hi-Bred International, Inc. | Functional expression of yeast nitrate transporter (ynt1) in maize to improve nitrate uptake |
| WO2011025860A1 (en) | 2009-08-28 | 2011-03-03 | E. I. Du Pont De Nemours And Company | Compositions and methods to control insect pests |
| EP2298915A1 (en) | 2004-06-30 | 2011-03-23 | Pioneer Hi-Bred International, Inc. | Methods of protecting plants from pathogenic fungi |
| WO2011036569A2 (en) | 2009-09-22 | 2011-03-31 | The Board Of Trustees Of The University Of Arkansas, N.A. | Rice cultivar cl 142-ar |
| WO2011041796A1 (en) | 2009-10-02 | 2011-04-07 | Pioneer Hi-Bred International, Inc. | Down-regulation of acc synthase for improved plant performance |
| EP2308986A1 (en) | 2006-05-17 | 2011-04-13 | Pioneer Hi-Bred International Inc. | Artificial plant minichromosomes |
| WO2011047191A1 (en) | 2009-10-15 | 2011-04-21 | The Board Of Trustees Of The University Of Arkansas, N.A. | Rice cultivar cl181-ar |
| US7935862B2 (en) | 2003-12-02 | 2011-05-03 | Syngenta Participations Ag | Targeted integration and stacking of DNA through homologous recombination |
| WO2011056544A1 (en) | 2009-10-26 | 2011-05-12 | Pioneer Hi-Bred International, Inc. | Somatic ovule specific promoter and methods of use |
| EP2322629A2 (en) | 2003-04-29 | 2011-05-18 | Pioneer Hi-Bred International Inc. | Novel glyphosate-n-acetyltransferase (GAT) genes |
| WO2011068567A1 (en) | 2009-07-10 | 2011-06-09 | Syngenta Participations Ag | Novel hydroxyphenylpyruvate dioxygenase polypeptides and methods of use |
| EP2333088A1 (en) | 2006-05-16 | 2011-06-15 | Pioneer Hi-Bred International, Inc. | Antifungal polypeptides |
| US7964774B2 (en) | 2008-05-14 | 2011-06-21 | Monsanto Technology Llc | Plants and seeds of spring canola variety SCV384196 |
| EP2338905A2 (en) | 2005-02-23 | 2011-06-29 | North Carolina State University | Alteration of tobacco alkaloid content through modification of specific cytochrome p450 genes |
| WO2011082318A2 (en) | 2009-12-30 | 2011-07-07 | Pioneer Hi-Bred International, Inc. | Methods and compositions for the introduction and regulated expression of genes in plants |
| WO2011082310A2 (en) | 2009-12-30 | 2011-07-07 | Pioneer Hi-Bred International, Inc. | Methods and compositions for targeted polynucleotide modification |
| WO2011082304A1 (en) | 2009-12-31 | 2011-07-07 | Pioneer Hi-Bred International, Inc. | Engineering plant resistance to diseases caused by pathogens |
| WO2011085062A1 (en) | 2010-01-06 | 2011-07-14 | Pioneer Hi-Bred International, Inc. | Identification of diurnal rhythms in photosynthetic and non-photosynthetic tissues from zea mays and use in improving crop plants |
| WO2011094199A1 (en) | 2010-01-26 | 2011-08-04 | Pioneer Hi-Bred International, Inc. | Polynucleotide and polypeptide sequences associated with herbicide tolerance |
| WO2011139431A1 (en) | 2010-05-06 | 2011-11-10 | Pioneer Hi-Bred International, Inc. | Maize acc synthase 3 gene and protein and uses thereof |
| WO2011163590A1 (en) | 2010-06-25 | 2011-12-29 | E. I. Du Pont De Nemours And Company | Compositions and methods for enhancing resistance to northern leaf blight in maize |
| EP2405013A2 (en) | 2008-01-17 | 2012-01-11 | Pioneer Hi-Bred International Inc. | Compositions and methods for the suppression of target polynucleotides from the family Aphididae |
| WO2012021785A1 (en) | 2010-08-13 | 2012-02-16 | Pioneer Hi-Bred International, Inc. | Compositions and methods comprising sequences having hydroxyphenylpyruvate dioxygenase (hppd) activity |
| WO2012027209A2 (en) | 2010-08-23 | 2012-03-01 | Pioneer Hi-Bred International, Inc. | Novel defensin variants and methods of use |
| WO2012071040A1 (en) | 2010-11-24 | 2012-05-31 | Pioneer Hi-Bred International, Inc. | Brassica gat event dp-073496-4 and compositions and methods for the identification and/or detection thereof |
| WO2012071039A1 (en) | 2010-11-24 | 2012-05-31 | Pioner Hi-Bred International, Inc. | Brassica gat event dp-061061-7 and compositions and methods for the identification and/or detection thereof |
| US8192984B2 (en) | 2001-09-04 | 2012-06-05 | Icon Genetics, Inc. | Creation of artificial internal ribosome entry site (IRES) elements |
| WO2012078949A2 (en) | 2010-12-09 | 2012-06-14 | Syngenta Participations Ag | Methods and compositions using small interfering rna (sirna) for nematode control in plants |
| WO2012082548A2 (en) | 2010-12-15 | 2012-06-21 | Syngenta Participations Ag | Soybean event syht0h2 and compositions and methods for detection thereof |
| WO2012092106A1 (en) | 2010-12-28 | 2012-07-05 | Pioneer Hi-Bred International, Inc. | Novel bacillus thuringiensis gene with lepidopteran activity |
| WO2012106321A1 (en) | 2011-02-01 | 2012-08-09 | Colorado Wheat Research Foundation, Inc. | Acetyl co-enzyme a carboxylase herbicide resistant plants |
| WO2012109515A1 (en) | 2011-02-11 | 2012-08-16 | Pioneer Hi-Bred International, Inc. | Synthetic insecticidal proteins active against corn rootworm |
| WO2012112411A1 (en) | 2011-02-15 | 2012-08-23 | Pioneer Hi-Bred International, Inc. | Root-preferred promoter and methods of use |
| US8257945B2 (en) | 2001-09-04 | 2012-09-04 | Icon Genetics, Inc. | Identification of eukaryotic internal ribosome entry site (IRES) elements |
| WO2012122369A1 (en) | 2011-03-10 | 2012-09-13 | Pioneer Hi-Bred International, Inc. | Novel bacillus thuringiensis gene with lepidopteran activity |
| WO2012129373A2 (en) | 2011-03-23 | 2012-09-27 | Pioneer Hi-Bred International, Inc. | Methods for producing a complex transgenic trait locus |
| WO2012131495A2 (en) | 2011-03-30 | 2012-10-04 | Universidad Nacional Autónoma de México | Mutant bacillus thuringiensis cry genes and methods of use |
| WO2012142311A1 (en) | 2011-04-15 | 2012-10-18 | Pioneer Hi-Bred International, Inc. | Self-reproducing hybrid plants |
| WO2012149470A1 (en) | 2011-04-27 | 2012-11-01 | Amyris, Inc. | Methods for genomic modification |
| WO2012148835A1 (en) | 2011-04-29 | 2012-11-01 | Pioneer Hi-Bred International, Inc. | Down-regulation of a homeodomain-leucine zipper i-class homeobox gene for improved plant performance |
| WO2012154824A1 (en) | 2011-05-09 | 2012-11-15 | E. I. Du Pont De Nemours And Company | Methods and compositions for silencing gene families using artificial micrornas |
| WO2012161982A1 (en) | 2011-05-23 | 2012-11-29 | E. I. Dupont De Nemours & Company | Chloroplast transit peptides and methods of their use |
| WO2012174139A2 (en) | 2011-06-14 | 2012-12-20 | Synthon Biopharmaceuticals B.V. | Compositions and methods for making and b ioc ont aining auxotrophic transgenic plants |
| EP2548964A2 (en) | 2008-01-17 | 2013-01-23 | Pioneer Hi-Bred International Inc. | Compositions and methods for the suppression of target polynucleotides from Lepidoptera |
| WO2013019411A1 (en) | 2011-08-03 | 2013-02-07 | E. I. Du Pont De Nemours And Company | Methods and compositions for targeted integration in a plant |
| WO2013033308A2 (en) | 2011-08-31 | 2013-03-07 | Pioneer Hi-Bred International, Inc. | Methods for tissue culture and transformation of sugarcane |
| EP2568048A1 (en) | 2007-06-29 | 2013-03-13 | Pioneer Hi-Bred International, Inc. | Methods for altering the genome of a monocot plant cell |
| EP2573183A1 (en) | 2009-01-22 | 2013-03-27 | Syngenta Participations AG. | Mutant hydroxyphenylpyruvate dioxgenase polypeptids and methods of use |
| WO2013063344A1 (en) | 2011-10-28 | 2013-05-02 | Pioneer Hi-Bred International, Inc. | Engineered pep carboxylase variants for improved plant productivity |
| WO2013063487A1 (en) | 2011-10-28 | 2013-05-02 | E. I. Du Pont De Nemours And Company | Methods and compositions for silencing genes using artificial micrornas |
| WO2013066423A2 (en) | 2011-06-21 | 2013-05-10 | Pioneer Hi-Bred International, Inc. | Methods and compositions for producing male sterile plants |
| WO2013066805A1 (en) | 2011-10-31 | 2013-05-10 | Pioneer Hi-Bred International, Inc. | Improving plant drought tolerance, nitrogen use efficiency and yield |
| US8445746B2 (en) | 2007-02-23 | 2013-05-21 | University Of Georgia Research Foundation, Inc. | Compositions and methods for identifying genetic sequences with toxin resistance in plants |
| WO2013096818A1 (en) | 2011-12-21 | 2013-06-27 | The Curators Of The University Of Missouri | Soybean variety s05-11268 |
| WO2013096810A1 (en) | 2011-12-21 | 2013-06-27 | The Curators Of The University Of Missouri | Soybean variety s05-11482 |
| WO2013103366A1 (en) | 2012-01-06 | 2013-07-11 | Pioneer Hi-Bred International, Inc. | A method to screen plants for genetic elements inducing parthenogenesis in plants |
| WO2013103371A1 (en) | 2012-01-06 | 2013-07-11 | Pioneer Hi-Bred International, Inc. | Ovule specific promoter and methods of use |
| WO2013103365A1 (en) | 2012-01-06 | 2013-07-11 | Pioneer Hi-Bred International, Inc. | Pollen preferred promoters and methods of use |
| WO2013104026A1 (en) | 2012-01-11 | 2013-07-18 | The Australian National University | Method for modulating plant root architecture |
| WO2013112686A1 (en) | 2012-01-27 | 2013-08-01 | Pioneer Hi-Bred International, Inc. | Methods and compositions for generating complex trait loci |
| WO2013122720A2 (en) | 2012-02-16 | 2013-08-22 | Syngenta Participations Ag | Engineered pesticidal proteins |
| WO2013138358A1 (en) | 2012-03-13 | 2013-09-19 | Pioneer Hi-Bred International, Inc. | Genetic reduction of male fertility in plants |
| WO2013138309A1 (en) | 2012-03-13 | 2013-09-19 | Pioneer Hi-Bred International, Inc. | Genetic reduction of male fertility in plants |
| WO2013166113A1 (en) | 2012-05-04 | 2013-11-07 | E. I. Du Pont De Nemours And Company | Compositions and methods comprising sequences having meganuclease activity |
| EP2664673A2 (en) | 2006-10-05 | 2013-11-20 | E. I. du Pont de Nemours and Company | Maize microRNA sequences |
| EP2666781A1 (en) | 2009-05-04 | 2013-11-27 | Pioneer Hi-Bred International, Inc. | Yield enhancement in plants by modulation of AP2 transcription factor |
| WO2013177400A1 (en) | 2012-05-24 | 2013-11-28 | Iowa State University Research Foundation, Inc. | Arabidopsis nonhost resistance gene(s) and use thereof to engineer disease resistant plants |
| WO2013188501A1 (en) | 2012-06-15 | 2013-12-19 | Pioneer Hi-Bred International, Inc. | Genetic loci associated with resistance of soybean to cyst nematode and methods of use |
| WO2013188291A2 (en) | 2012-06-15 | 2013-12-19 | E. I. Du Pont De Nemours And Company | Methods and compositions involving als variants with native substrate preference |
| WO2013192256A1 (en) | 2012-06-22 | 2013-12-27 | Syngenta Participations Ag | Biological control of coleopteran pests |
| WO2014059155A1 (en) | 2012-10-11 | 2014-04-17 | Pioneer Hi-Bred International, Inc. | Guard cell promoters and uses thereof |
| WO2014062544A2 (en) | 2012-10-15 | 2014-04-24 | Pioneer Hi-Bred International, Inc. | Methods and compositions to enhance activity of cry endotoxins |
| EP2730587A2 (en) | 2006-02-09 | 2014-05-14 | Pioneer Hi-Bred International, Inc. | Genes for enhancing nitrogen utilization efficiency in crop plants |
| WO2014081673A2 (en) | 2012-11-20 | 2014-05-30 | Pioneer Hi-Bred International, Inc. | Engineering plants for efficient uptake and utilization of urea to improve crop production |
| US8754293B2 (en) | 2011-09-09 | 2014-06-17 | Nunhems B.V. | Lettuce variety intred |
| WO2014093485A1 (en) | 2012-12-13 | 2014-06-19 | Pioneer Hi-Bred International, Inc. | Methods and compositions for producing and selecting transgenic plants |
| WO2014100525A2 (en) | 2012-12-21 | 2014-06-26 | Pioneer Hi-Bred International, Inc. | Compositions and methods for auxin-analog conjugation |
| US8785729B2 (en) | 2011-08-09 | 2014-07-22 | Nunhems, B.V. | Lettuce variety redglace |
| WO2014116989A1 (en) | 2013-01-25 | 2014-07-31 | Pioneer Hi-Bred International, Inc. | Maize event dp-032218-9 and methods for detection thereof |
| US8835720B2 (en) | 2012-04-26 | 2014-09-16 | Monsanto Technology Llc | Plants and seeds of spring canola variety SCV967592 |
| WO2014143996A2 (en) | 2013-03-15 | 2014-09-18 | Pioneer Hi-Bred International, Inc. | Compositions and methods of use of acc oxidase polynucleotides and polypeptides |
| WO2014153234A1 (en) | 2013-03-14 | 2014-09-25 | Pioneer Hi-Bred International, Inc. | Compositions having dicamba decarboxylase activity and methods of use |
| WO2014152507A2 (en) | 2013-03-15 | 2014-09-25 | Pioneer Hi-Bred International, Inc. | Modulation of acc deaminase expression |
| WO2014150914A2 (en) | 2013-03-15 | 2014-09-25 | Pioneer Hi-Bred International, Inc. | Phi-4 polypeptides and methods for their use |
| WO2014153242A1 (en) | 2013-03-14 | 2014-09-25 | Pioneer Hi-Bred International, Inc. | Compositions having dicamba decarboxylase activity and methods of use |
| WO2014153254A2 (en) | 2013-03-14 | 2014-09-25 | Pioneer Hi-Bred International Inc. | Compositions and methods to control insect pests |
| WO2014160383A1 (en) | 2013-03-13 | 2014-10-02 | E. I. Dupont De Nemours & Company | Production of small interfering rnas in planta |
| WO2014159306A1 (en) | 2013-03-13 | 2014-10-02 | Pioneer Hi-Bred International, Inc. | Glyphosate application for weed control in brassica |
| WO2014160122A1 (en) | 2013-03-14 | 2014-10-02 | Pioneer Hi-Bred International, Inc. | Maize stress related transcription factor 18 and uses thereof |
| WO2014159113A1 (en) | 2013-03-12 | 2014-10-02 | Pioneer Hi-Bred International, Inc. | Root-preferred promoter and methods of use |
| WO2014164116A1 (en) | 2013-03-13 | 2014-10-09 | Pioneer Hi-Bred International, Inc. | Functional expression of bacterial major facilitator superfamily (sfm) gene in maize to improve agronomic traits and grain yield |
| WO2014164074A1 (en) | 2013-03-13 | 2014-10-09 | Pioneer Hi-Bred International, Inc. | Enhanced nitrate uptake and nitrate translocation by over-expressing maize functional low-affinity nitrate transporters in transgenic maize |
| WO2014164399A1 (en) | 2013-03-12 | 2014-10-09 | Pioneer Hi-Bred International, Inc. | Root-preferred promoter and methods of use |
| WO2014164466A1 (en) | 2013-03-12 | 2014-10-09 | E. I. Du Pont De Nemours And Company | Methods for the identification of variant recognition sites for rare-cutting engineered double-strand-break-inducing agents and compositions and uses thereof |
| WO2014164828A2 (en) | 2013-03-11 | 2014-10-09 | Pioneer Hi-Bred International, Inc. | Methods and compositions employing a sulfonylurea-dependent stabilization domain |
| WO2014164961A2 (en) | 2013-03-12 | 2014-10-09 | Pioneer Hi-Bred International, Inc. | Manipulation of dominant male sterility |
| WO2014164014A1 (en) | 2013-03-11 | 2014-10-09 | Pioneer Hi-Bred International, Inc. | Genes for improving nutrient uptake and abiotic stress tolerance in plants |
| WO2014164775A1 (en) | 2013-03-11 | 2014-10-09 | Pioneer Hi-Bred International, Inc. | Methods and compositions to improve the spread of chemical signals in plants |
| US8859857B2 (en) | 2012-04-26 | 2014-10-14 | Monsanto Technology Llc | Plants and seeds of spring canola variety SCV259778 |
| US8878009B2 (en) | 2012-04-26 | 2014-11-04 | Monsanto Technology, LLP | Plants and seeds of spring canola variety SCV318181 |
| WO2014201511A1 (en) | 2013-06-21 | 2014-12-24 | Gary David Housley | Method and apparatus for close-field electroporation |
| EP2821490A2 (en) | 2008-10-30 | 2015-01-07 | Pioneer Hi-Bred International Inc. | Manipulation of glutamine synthetases (GS) to improve nitrogen use efficiency and grain yield in higher plants |
| WO2015006105A1 (en) | 2013-07-09 | 2015-01-15 | Board Of Trustees Of Michigan State University | Transgenic plants produced with a k-domain, and methods and expression cassettes related thereto |
| WO2015017510A1 (en) | 2013-07-31 | 2015-02-05 | E. I. Du Pont De Nemours And Company | Modification of soybean seed composition to enhance feed, food and other industrial applications of soybean products |
| WO2015021139A2 (en) | 2013-08-08 | 2015-02-12 | Pioneer Hi-Bred International, Inc. | Insecticidal polypeptides having broad spectrum activity and uses thereof |
| WO2015023846A2 (en) | 2013-08-16 | 2015-02-19 | Pioneer Hi-Bred International, Inc. | Insecticidal proteins and methods for their use |
| WO2015038622A1 (en) | 2013-09-11 | 2015-03-19 | Pioneer Hi-Bred International, Inc. | Plant regulatory elements and methods of use thereof |
| WO2015038734A2 (en) | 2013-09-13 | 2015-03-19 | Pioneer Hi-Bred International, Inc. | Insecticidal proteins and methods for their use |
| WO2015057600A1 (en) | 2013-10-18 | 2015-04-23 | E. I. Du Pont De Nemours And Company | Glyphosate-n-acetyltransferase (glyat) sequences and methods of use |
| WO2015066011A2 (en) | 2013-10-29 | 2015-05-07 | Pioneer Hi-Bred International, Inc. | Self-reproducing hybrid plants |
| WO2015095804A1 (en) | 2013-12-19 | 2015-06-25 | Amyris, Inc. | Methods for genomic integration |
| WO2015120270A1 (en) | 2014-02-07 | 2015-08-13 | Pioneer Hi Bred International, Inc. | Insecticidal proteins and methods for their use |
| WO2015120276A1 (en) | 2014-02-07 | 2015-08-13 | Pioneer Hi Bred International Inc | Insecticidal proteins and methods for their use |
| WO2015164457A1 (en) | 2014-04-22 | 2015-10-29 | E. I. Du Pont De Nemours And Company | Plastidic carbonic anhydrase genes for oil augmentation in seeds with increased dgat expression |
| WO2016000647A1 (en) | 2014-07-03 | 2016-01-07 | Pioneer Overseas Corporation | Plants having enhanced tolerance to insect pests and related constructs and methods involving insect tolerance genes |
| WO2016022516A1 (en) | 2014-08-08 | 2016-02-11 | Pioneer Hi-Bred International, Inc. | Ubiquitin promoters and introns and methods of use |
| WO2016040030A1 (en) | 2014-09-12 | 2016-03-17 | E. I. Du Pont De Nemours And Company | Generation of site-specific-integration sites for complex trait loci in corn and soybean, and methods of use |
| US9290777B2 (en) | 2007-02-05 | 2016-03-22 | National University Of Singapore | Putative cytokinin receptor and methods for use thereof |
| WO2016044092A1 (en) | 2014-09-17 | 2016-03-24 | Pioneer Hi Bred International Inc | Compositions and methods to control insect pests |
| WO2016060949A1 (en) | 2014-10-16 | 2016-04-21 | Pioneer Hi-Bred International, Inc. | Insecticidal polypeptides having broad spectrum activity and uses thereof |
| WO2016061206A1 (en) | 2014-10-16 | 2016-04-21 | Pioneer Hi-Bred International, Inc. | Insecticidal proteins and methods for their use |
| WO2016061197A1 (en) | 2014-10-16 | 2016-04-21 | Pioneer Hi-Bred International, Inc. | Insecticidal polypeptides having improved activity spectrum and uses thereof |
| WO2016094165A1 (en) | 2014-12-12 | 2016-06-16 | Syngenta Participations Ag | Compositions and methods for controlling plant pests |
| WO2016100309A1 (en) | 2014-12-16 | 2016-06-23 | Pioneer Hi-Bred International, Inc. | Restoration of male fertility in wheat |
| WO2016100333A1 (en) | 2014-12-15 | 2016-06-23 | Syngenta Participations Ag | Pesticidal microrna carriers and use thereof |
| WO2016099916A1 (en) | 2014-12-19 | 2016-06-23 | E. I. Du Pont De Nemours And Company | Polylactic acid compositions with accelerated degradation rate and increased heat stability |
| WO2016105696A1 (en) | 2014-12-23 | 2016-06-30 | Syngenta Participations Ag | Biological control of coleopteran pests |
| US9380756B2 (en) | 2012-01-04 | 2016-07-05 | Nunhems B.V. | Lettuce variety multigreen 50 |
| WO2016144688A1 (en) | 2015-03-11 | 2016-09-15 | Pioneer Hi Bred International Inc | Insecticidal combinations of pip-72 and methods of use |
| WO2016149352A1 (en) | 2015-03-19 | 2016-09-22 | Pioneer Hi-Bred International Inc | Methods and compositions for accelerated trait introgression |
| EP3091076A1 (en) | 2015-05-07 | 2016-11-09 | Limagrain Europe | Polynucleotide responsible of haploid induction in maize plants and related processes |
| WO2016186986A1 (en) | 2015-05-19 | 2016-11-24 | Pioneer Hi Bred International Inc | Insecticidal proteins and methods for their use |
| WO2016205445A1 (en) | 2015-06-16 | 2016-12-22 | Pioneer Hi-Bred International, Inc. | Compositions and methods to control insect pests |
| WO2017023486A1 (en) | 2015-08-06 | 2017-02-09 | Pioneer Hi-Bred International, Inc. | Plant derived insecticidal proteins and methods for their use |
| WO2017040343A1 (en) | 2015-08-28 | 2017-03-09 | Pioneer Hi-Bred International, Inc. | Ochrobactrum-mediated transformation of plants |
| WO2017066597A1 (en) | 2015-10-16 | 2017-04-20 | Pioneer Hi-Bred International, Inc. | Generating maize plants with enhanced resistance to northern leaf blight |
| WO2017074547A1 (en) | 2015-10-30 | 2017-05-04 | Pioneer Hi-Bred International, Inc. | Methods and compositions for rapid plant transformation |
| WO2017079026A1 (en) | 2015-11-06 | 2017-05-11 | E. I. Du Pont De Nemours And Company | Generation of complex trait loci in soybean and methods of use |
| WO2017078836A1 (en) | 2015-11-06 | 2017-05-11 | Pioneer Hi-Bred International, Inc. | Methods and compositions of improved plant transformation |
| WO2017112006A1 (en) | 2015-12-22 | 2017-06-29 | Pioneer Hi-Bred International, Inc. | Tissue-preferred promoters and methods of use |
| WO2017161264A1 (en) | 2016-03-18 | 2017-09-21 | Pioneer Hi-Bred International, Inc. | Methods and compositions for producing clonal, non-reduced, non-recombined gametes |
| WO2017180715A2 (en) | 2016-04-14 | 2017-10-19 | Pioneer Hi-Bred International, Inc. | Insecticidal polypeptides having improved activity spectrum and uses thereof |
| WO2017184673A1 (en) | 2016-04-19 | 2017-10-26 | Pioneer Hi-Bred International, Inc. | Insecticidal combinations of polypeptides having improved activity spectrum and uses thereof |
| WO2017192560A1 (en) | 2016-05-04 | 2017-11-09 | Pioneer Hi-Bred International, Inc. | Insecticidal proteins and methods for their use |
| US9816102B2 (en) | 2012-09-13 | 2017-11-14 | Indiana University Research And Technology Corporation | Compositions and systems for conferring disease resistance in plants and methods of use thereof |
| WO2017218207A1 (en) | 2016-06-16 | 2017-12-21 | Pioneer Hi-Bred International, Inc. | Compositions and methods to control insect pests |
| WO2017222821A2 (en) | 2016-06-24 | 2017-12-28 | Pioneer Hi-Bred International, Inc. | Plant regulatory elements and methods of use thereof |
| WO2018005411A1 (en) | 2016-07-01 | 2018-01-04 | Pioneer Hi-Bred International, Inc. | Insecticidal proteins from plants and methods for their use |
| WO2018013333A1 (en) | 2016-07-12 | 2018-01-18 | Pioneer Hi-Bred International, Inc. | Compositions and methods to control insect pests |
| WO2018026774A1 (en) | 2016-08-05 | 2018-02-08 | Syngenta Participations Ag | Control of coleopteran pests using rna molecules |
| WO2018078390A1 (en) | 2016-10-31 | 2018-05-03 | Institute Of Genetics And Developmental Biology Chinese Academy Of Sciences | Compositions and methods for enhancing abiotic stress tolerance |
| WO2018084936A1 (en) | 2016-11-01 | 2018-05-11 | Pioneer Hi-Bred International, Inc. | Insecticidal proteins and methods for their use |
| WO2018111551A1 (en) | 2016-12-14 | 2018-06-21 | Pioneer Hi-Bred International, Inc. | Insecticidal proteins and methods for their use |
| WO2018118811A1 (en) | 2016-12-22 | 2018-06-28 | Pioneer Hi-Bred International, Inc. | Insecticidal proteins and methods for their use |
| WO2018140214A1 (en) | 2017-01-24 | 2018-08-02 | Pioneer Hi-Bred International, Inc. | Nematicidal protein from pseudomonas |
| WO2018148001A1 (en) | 2017-02-08 | 2018-08-16 | Pioneer Hi-Bred International Inc | Insecticidal combinations of plant derived insecticidal proteins and methods for their use |
| WO2018208882A1 (en) | 2017-05-11 | 2018-11-15 | Pioneer Hi-Bred International, Inc. | Insecticidal proteins and methods for their use |
| WO2018217333A1 (en) | 2017-05-26 | 2018-11-29 | Pioneer Hi-Bred International, Inc. | Insecticidal polypeptides having improved activity spectrum and uses thereof |
| EP3441470A1 (en) | 2008-09-26 | 2019-02-13 | BASF Agrochemical Products, B.V. | Herbicide-resistant ahas-mutants and methods of use |
| WO2019049111A1 (en) | 2017-09-11 | 2019-03-14 | R. J. Reynolds Tobacco Company | Methods and compositions for increasing expression of genes of interest in a plant by co-expression with p21 |
| WO2019060383A1 (en) | 2017-09-25 | 2019-03-28 | Pioneer Hi-Bred, International, Inc. | Tissue-preferred promoters and methods of use |
| WO2019074598A1 (en) | 2017-10-13 | 2019-04-18 | Pioneer Hi-Bred International, Inc. | Virus-induced gene silencing technology for insect control in maize |
| WO2019076891A2 (en) | 2017-10-18 | 2019-04-25 | Syngenta Participations Ag | Control of hemipteran pests using rna molecules |
| WO2019162163A1 (en) | 2018-02-26 | 2019-08-29 | Devgen Nv | Control of insect pests using rna molecules |
| WO2019169150A1 (en) | 2018-03-02 | 2019-09-06 | Pioneer Hi-Bred International, Inc. | Plant health assay |
| US10428336B2 (en) | 2013-10-16 | 2019-10-01 | The Australian National University | Method for modulating plant growth |
| WO2019206780A1 (en) | 2018-04-27 | 2019-10-31 | Devgen Nv | Control of insect pests using rna molecules |
| WO2019226508A1 (en) | 2018-05-22 | 2019-11-28 | Pioneer Hi-Bred International, Inc. | Plant regulatory elements and methods of use thereof |
| WO2020005933A1 (en) | 2018-06-28 | 2020-01-02 | Pioneer Hi-Bred International, Inc. | Methods for selecting transformed plants |
| WO2020046701A1 (en) | 2018-08-29 | 2020-03-05 | Pioneer Hi-Bred International, Inc. | Insecticidal proteins and methods for their use |
| WO2020092487A1 (en) | 2018-10-31 | 2020-05-07 | Pioneer Hi-Bred International, Inc. | Compositions and methods for ochrobactrum-mediated plant transformation |
| WO2020185751A1 (en) | 2019-03-11 | 2020-09-17 | Pioneer Hi-Bred International, Inc. | Methods for clonal plant production |
| WO2020187798A1 (en) | 2019-03-21 | 2020-09-24 | Devgen Nv | Control of insect pests using rna molecules |
| WO2020198408A1 (en) | 2019-03-27 | 2020-10-01 | Pioneer Hi-Bred International, Inc. | Plant explant transformation |
| WO2020198496A1 (en) | 2019-03-28 | 2020-10-01 | Pioneer Hi-Bred International, Inc. | Modified agrobacterium strains and use thereof for plant transformation |
| WO2020214986A1 (en) | 2019-04-18 | 2020-10-22 | Pioneer Hi-Bred International, Inc. | Embryogenesis factors for cellular reprogramming of a plant cell |
| WO2021158455A1 (en) | 2020-02-04 | 2021-08-12 | Dow Agrosciences Llc | Compositions having pesticidal utility and processes related thereto |
| US11140902B2 (en) | 2016-09-27 | 2021-10-12 | University Of Florida Research Foundation, Inc. | Insect toxin delivery mediated by a densovirus coat protein |
| WO2021252238A1 (en) | 2020-06-12 | 2021-12-16 | Pioneer Hi-Bred International, Inc. | Alteration of seed composition in plants |
| WO2021257206A1 (en) | 2020-06-17 | 2021-12-23 | Pioneer Hi-Bred International, Inc. | Generating maize plants with enhanced resistance to northern leaf blight |
| WO2022015619A2 (en) | 2020-07-14 | 2022-01-20 | Pioneer Hi-Bred International, Inc. | Insecticidal proteins and methods for their use |
| WO2022035653A2 (en) | 2020-08-10 | 2022-02-17 | E. I. Du Pont De Nemours And Company | Compositions and methods for enhancing resistance to northern leaf blight in maize |
| WO2022035649A1 (en) | 2020-08-10 | 2022-02-17 | Pioneer Hi-Bred International, Inc. | Plant regulatory elements and methods of use thereof |
| WO2022072335A2 (en) | 2020-09-30 | 2022-04-07 | Pioneer Hi-Bred International, Inc. | Rapid transformation of monocot leaf explants |
| WO2022087616A1 (en) | 2020-10-21 | 2022-04-28 | Pioneer Hi-Bred International, Inc. | Parthenogenesis factors and methods of using same |
| WO2022115524A2 (en) | 2020-11-24 | 2022-06-02 | AgBiome, Inc. | Pesticidal genes and methods of use |
| US11459579B2 (en) | 2013-07-09 | 2022-10-04 | Board Of Trustees Of Michigan State University | Transgenic plants produced with a K-domain, and methods and expression cassettes related thereto |
| EP4079857A2 (en) | 2016-08-05 | 2022-10-26 | Syngenta Participations Ag | Control of coleopteran pests using rna molecules |
| WO2022226316A1 (en) | 2021-04-22 | 2022-10-27 | Precision Biosciences, Inc. | Compositions and methods for generating male sterile plants |
| WO2022236060A1 (en) | 2021-05-06 | 2022-11-10 | AgBiome, Inc. | Pesticidal genes and methods of use |
| EP4122947A1 (en) | 2017-12-19 | 2023-01-25 | Pioneer Hi-Bred International, Inc. | Insecticidal polypeptides and uses thereof |
| WO2023012342A1 (en) | 2021-08-06 | 2023-02-09 | Kws Vegetables B.V. | Durable downy mildew resistance in spinach |
| WO2023107943A1 (en) | 2021-12-07 | 2023-06-15 | AgBiome, Inc. | Pesticidal genes and methods of use |
| EP4219731A2 (en) | 2016-05-18 | 2023-08-02 | Amyris, Inc. | Compositions and methods for genomic integration of nucleic acids into exogenous landing pads |
| WO2023183918A1 (en) | 2022-03-25 | 2023-09-28 | Pioneer Hi-Bred International, Inc. | Methods of parthenogenic haploid induction and haploid chromosome doubling |
| WO2023224815A2 (en) | 2022-05-18 | 2023-11-23 | Corteva Agriscience Llc | Compositions having pesticidal utility and processes related thereto |
| WO2024023578A1 (en) | 2022-07-28 | 2024-02-01 | Institut Pasteur | Hsc70-4 in host-induced and spray-induced gene silencing |
| US11895964B2 (en) | 2021-06-02 | 2024-02-13 | Nutrien Ag Solutions, Inc. | Inbred rice line DG263L |
| US11905518B2 (en) | 2018-02-12 | 2024-02-20 | Curators Of The University Of Missouri | Small auxin upregulated (SAUR) gene for the improvement of root system architecture, waterlogging tolerance, drought resistance and yield in plants and methods of uses |
| WO2024044596A1 (en) | 2022-08-23 | 2024-02-29 | AgBiome, Inc. | Pesticidal genes and methods of use |
Families Citing this family (89)
| Publication number | Priority date | Publication date | Assignee | Title |
|---|---|---|---|---|
| US7227057B2 (en) | 1997-06-03 | 2007-06-05 | Chromatin, Inc. | Plant centromere compositions |
| US7119250B2 (en) * | 1997-06-03 | 2006-10-10 | The University Of Chicago | Plant centromere compositions |
| US7235716B2 (en) | 1997-06-03 | 2007-06-26 | Chromatin, Inc. | Plant centromere compositions |
| US7193128B2 (en) * | 1997-06-03 | 2007-03-20 | Chromatin, Inc. | Methods for generating or increasing revenues from crops |
| US6632980B1 (en) | 1997-10-24 | 2003-10-14 | E. I. Du Pont De Nemours And Company | Binary viral expression system in plants |
| US7102055B1 (en) * | 1997-11-18 | 2006-09-05 | Pioneer Hi-Bred International, Inc. | Compositions and methods for the targeted insertion of a nucleotide sequence of interest into the genome of a plant |
| WO1999025840A1 (en) | 1997-11-18 | 1999-05-27 | Pioneer Hi-Bred International, Inc. | A novel method for the integration of foreign dna into eukaryoticgenomes |
| DE69831265T2 (en) * | 1997-11-18 | 2006-06-08 | Pioneer Hi-Bred International, Inc. | COMPOSITIONS AND METHODS FOR THE GENETIC MODIFICATION OF PLANTS |
| WO1999025855A1 (en) | 1997-11-18 | 1999-05-27 | Pioneer Hi-Bred International, Inc. | Mobilization of viral genomes from t-dna using site-specific recombination systems |
| US7989202B1 (en) | 1999-03-18 | 2011-08-02 | The University Of Chicago | Plant centromere compositions |
| US7126041B1 (en) * | 1999-12-10 | 2006-10-24 | North Carolina State Unversity | High efficiency gene targeting in plants |
| US20040231006A1 (en) * | 2000-04-12 | 2004-11-18 | Silver Daniel P. | Self-extinguishing recombinases, nucleic acids encoding them and methods of using the same |
| US7560622B2 (en) * | 2000-10-06 | 2009-07-14 | Pioneer Hi-Bred International, Inc. | Methods and compositions relating to the generation of partially transgenic organisms |
| NZ529597A (en) * | 2001-05-30 | 2005-12-23 | Chromos Molecular Systems Inc | Plant artificial chromosomes, uses thereof and methods of preparing plant artificial chromosomes |
| US20030082591A1 (en) * | 2001-07-24 | 2003-05-01 | Donald Awrey | Methods for gene disruption and uses thereof |
| EP1428885A4 (en) * | 2001-08-28 | 2006-08-09 | Japan Tobacco Inc | Method of modifying genome in higher plant |
| DE10143205A1 (en) * | 2001-09-04 | 2003-03-20 | Icon Genetics Ag | Process for protein production in plants |
| US20050158291A1 (en) * | 2002-02-21 | 2005-07-21 | Lionel Breton | Pet food composition for skin photoprotection |
| US9155544B2 (en) | 2002-03-20 | 2015-10-13 | P Tech, Llc | Robotic systems and methods |
| US8304233B2 (en) | 2002-06-04 | 2012-11-06 | Poetic Genetics, Llc | Methods of unidirectional, site-specific integration into a genome, compositions and kits for practicing the same |
| US7365186B2 (en) * | 2002-11-22 | 2008-04-29 | Arborgen, Llc | Vascular-preferred promoter sequences and uses thereof |
| US20040137624A1 (en) * | 2002-12-27 | 2004-07-15 | Lowe Brenda A. | Methods of site-directed transformation |
| DE10354616A1 (en) * | 2003-11-21 | 2005-06-23 | Degussa Ag | rubber compounds |
| EP2434020B1 (en) | 2004-01-22 | 2014-11-05 | Dnavec Research Inc. | Method of producing Paramyxoviridae viral vectors using a hybrid promoter comprising a cytomegalovirus enhancer and a chicken beta-actin promoter |
| EP1718754A1 (en) * | 2004-02-23 | 2006-11-08 | Chromatin, Inc. | Plants modified with mini-chromosomes |
| US20090217400A1 (en) * | 2004-02-26 | 2009-08-27 | State Of Israel, Ministry Of Agriculture, Agricultural Research Organization | Enzymes, cells and methods for site specific recombination at asymmetric sites |
| US7319403B2 (en) * | 2004-03-08 | 2008-01-15 | Noel Woodard | Combination carbon monoxide and wireless E-911 location alarm |
| US20070197474A1 (en) * | 2004-03-30 | 2007-08-23 | Clinton William P | Methods for controlling plants pathogens using N-phosphonomethylglycine |
| WO2005115130A2 (en) * | 2004-05-24 | 2005-12-08 | The State Of Israel - Ministry Of Agriculture & Rural Development, Agricultural Research Organization | Transgenic ficus, method for producing same and use thereof |
| KR100579836B1 (en) * | 2004-06-04 | 2006-05-15 | 삼성전자주식회사 | Preparation method of toner having micro radius |
| BRPI0513706A (en) | 2004-07-20 | 2008-05-13 | Symphogen As | recombinant polyclonal anti-rhesus antibody and production methods |
| US7453025B2 (en) | 2004-09-22 | 2008-11-18 | Arborgen, Llc | Reproductive ablation constructs |
| PT1885176T (en) * | 2005-05-27 | 2016-11-28 | Monsanto Technology Llc | Soybean event mon89788 and methods for detection thereof |
| WO2007030510A2 (en) * | 2005-09-08 | 2007-03-15 | Chromatin Inc. | Plants modified with mini-chromosomes |
| JP2009524406A (en) * | 2005-10-13 | 2009-07-02 | ビーシー キャンサー エージェンシー | Modular genomes for synthetic biology and metabolic engineering |
| US9157066B2 (en) | 2005-12-13 | 2015-10-13 | The Trustees Of The University Of Pennsylvania | Transcriptome transfer produces cellular phenotype conversion |
| ES2433669T3 (en) | 2005-12-13 | 2013-12-12 | The Trustees Of The University Of Pennsylvania | Methods for phototransfecting nucleic acid in living cells |
| US10647960B2 (en) * | 2005-12-13 | 2020-05-12 | The Trustees Of The University Of Pennsylvania | Transcriptome transfer produces cellular phenotype conversion |
| US20070143881A1 (en) * | 2005-12-16 | 2007-06-21 | Pioneer Hi-Bred International, Inc. | Methods and Compositions for Improving the Efficiency of Site-Specific Polynucleotide Exchange |
| US8058509B2 (en) * | 2005-12-21 | 2011-11-15 | Pioneer Hi-Bred International, Inc. | Methods and compositions for in planta production of inverted repeats |
| CN105002207A (en) | 2006-05-09 | 2015-10-28 | 密苏里大学学监 | Plant artificial chromosome platform via telomere truncation |
| CA3014139A1 (en) | 2006-05-12 | 2007-11-22 | Monsanto Technology Llc | T-dna constructs for obtaining marker-free transgenic plants, and plants comprising such constructs |
| UA106193C2 (en) * | 2006-08-11 | 2014-08-11 | ДАУ АГРОСАЙЕНСИЗ ЕлЕлСи | Homological recombination mediated by zinc finger nucleases |
| US8614089B2 (en) * | 2007-03-15 | 2013-12-24 | Chromatin, Inc. | Centromere sequences and minichromosomes |
| WO2008130985A2 (en) * | 2007-04-16 | 2008-10-30 | Monsanto Technology, Llc | Plants with multiple transgenes on a chromosome |
| US20100162428A1 (en) * | 2007-05-31 | 2010-06-24 | Basf Plant Science Gmbh | Method Of Excising A Nucleic Acid Sequence From A Plant Genome |
| DE102007027595A1 (en) | 2007-06-12 | 2008-12-18 | Henkel Ag & Co. Kgaa | adhesive compositions |
| US8450106B2 (en) * | 2007-10-17 | 2013-05-28 | The Ohio State University Research Foundation | Oncolytic virus |
| US8125907B2 (en) * | 2008-06-12 | 2012-02-28 | Talari Networks Incorporated | Flow-based adaptive private network with multiple WAN-paths |
| WO2010111698A2 (en) * | 2009-03-27 | 2010-09-30 | E. I. Du Pont De Nemours And Company | Dielectric heat-transfer fluid |
| WO2010118045A1 (en) * | 2009-04-07 | 2010-10-14 | The Trustees Of The University Of Pennsylvania | Transcriptome transfer produces cellular phenotype conversion |
| WO2010135324A1 (en) | 2009-05-18 | 2010-11-25 | Monsanto Technology Llc | Use of glyphosate for disease suppression and yield enhancement in soybean |
| US9813315B2 (en) | 2009-06-11 | 2017-11-07 | Talari Networks Incorporated | Methods and apparatus for providing adaptive private network centralized management system time correlated playback of network traffic |
| PT3241435T (en) | 2009-07-08 | 2021-08-26 | Kymab Ltd | Animal models and therapeutic molecules |
| US20120204278A1 (en) | 2009-07-08 | 2012-08-09 | Kymab Limited | Animal models and therapeutic molecules |
| US9445581B2 (en) | 2012-03-28 | 2016-09-20 | Kymab Limited | Animal models and therapeutic molecules |
| EP2456894A4 (en) | 2009-07-23 | 2013-09-25 | Chromatin Inc | Sorghum centromere sequences and minichromosomes |
| WO2011050271A1 (en) * | 2009-10-23 | 2011-04-28 | Monsanto Technology Llc | Methods and compositions for expression of transgenes in plants |
| US9796788B2 (en) | 2010-02-08 | 2017-10-24 | Regeneron Pharmaceuticals, Inc. | Mice expressing a limited immunoglobulin light chain repertoire |
| US20130045492A1 (en) | 2010-02-08 | 2013-02-21 | Regeneron Pharmaceuticals, Inc. | Methods For Making Fully Human Bispecific Antibodies Using A Common Light Chain |
| EP2501817B2 (en) | 2010-02-08 | 2021-04-21 | Regeneron Pharmaceuticals, Inc. | Common light chain mouse |
| CN101886074B (en) * | 2010-06-03 | 2012-05-02 | 中国农业科学院生物技术研究所 | GhPsbP promoter high-effectively expressed by cotton chlorenchyma |
| US8878548B2 (en) | 2010-06-11 | 2014-11-04 | Baker Hughes Incorporated | Method for treating and sealing piezoelectric tuning forks |
| EP2471909A1 (en) | 2010-12-30 | 2012-07-04 | SIRION BIOTECH GmbH | Nucleic acid molecules for generating adenoviral vectors |
| SG2014007686A (en) | 2011-08-05 | 2014-03-28 | Regeneron Pharma | Humanized universal light chain mice |
| EP3839049A3 (en) | 2011-09-19 | 2021-10-20 | Kymab Limited | Antibodies, variable domains & chains tailored for human use |
| EP2761008A1 (en) | 2011-09-26 | 2014-08-06 | Kymab Limited | Chimaeric surrogate light chains (slc) comprising human vpreb |
| US9253965B2 (en) | 2012-03-28 | 2016-02-09 | Kymab Limited | Animal models and therapeutic molecules |
| US10195581B2 (en) | 2012-03-26 | 2019-02-05 | Unchained Labs | Parallel reactor systems and methods for preparing materials |
| GB2502127A (en) | 2012-05-17 | 2013-11-20 | Kymab Ltd | Multivalent antibodies and in vivo methods for their production |
| US10251377B2 (en) | 2012-03-28 | 2019-04-09 | Kymab Limited | Transgenic non-human vertebrate for the expression of class-switched, fully human, antibodies |
| US9788534B2 (en) | 2013-03-18 | 2017-10-17 | Kymab Limited | Animal models and therapeutic molecules |
| EP2992335B1 (en) | 2013-04-30 | 2018-04-18 | Unchained Labs | Methods for sampling from non-atmospheric vessels in a parallel reactor system |
| US11707056B2 (en) | 2013-05-02 | 2023-07-25 | Kymab Limited | Animals, repertoires and methods |
| US9783593B2 (en) | 2013-05-02 | 2017-10-10 | Kymab Limited | Antibodies, variable domains and chains tailored for human use |
| WO2015049517A2 (en) | 2013-10-01 | 2015-04-09 | Kymab Limited | Animal models and therapeutic molecules |
| KR102601491B1 (en) | 2014-03-21 | 2023-11-13 | 리제너론 파마슈티칼스 인코포레이티드 | Non-human animals that make single domain binding proteins |
| CN113337533A (en) | 2014-12-23 | 2021-09-03 | 先正达参股股份有限公司 | Methods and compositions for identifying and enriching cells comprising site-specific genomic modifications |
| US10439908B2 (en) | 2014-12-23 | 2019-10-08 | Talari Networks Incorporated | Methods and apparatus for providing adaptive private network centralized management system time correlated playback of network traffic |
| WO2016119703A1 (en) | 2015-01-27 | 2016-08-04 | 中国科学院遗传与发育生物学研究所 | Method for conducting site-specific modification on entire plant via gene transient expression |
| CA2975709A1 (en) * | 2015-02-02 | 2016-08-11 | Cellectis | Agrobacterium-mediated genome modification without t-dna integration |
| US11111314B2 (en) | 2015-03-19 | 2021-09-07 | Regeneron Pharmaceuticals, Inc. | Non-human animals that select for light chain variable regions that bind antigen |
| EP3095870A1 (en) | 2015-05-19 | 2016-11-23 | Kws Saat Se | Methods for the in planta transformation of plants and manufacturing processes and products based and obtainable therefrom |
| CA2994883A1 (en) | 2015-08-14 | 2017-02-23 | Institute Of Genetics And Developmental Biology, Chinese Academy Of Scnces | Method for obtaining glyphosate-resistant rice by site-directed nucleotide substitution |
| US10372834B2 (en) * | 2016-01-15 | 2019-08-06 | DISCUS Software Company | Creating and using an integrated technical data package |
| JP7402503B2 (en) | 2017-06-14 | 2023-12-21 | テクニスチェ ユニベルシタト ドレスデン | Methods and means for genetically modifying the genome using designer DNA recombinase |
| MX2022011408A (en) | 2020-03-15 | 2022-12-15 | Proteinea Inc | Recombinant protein production in insects. |
| CA3197678A1 (en) | 2020-10-21 | 2022-04-28 | Pioneer Hi-Bred International, Inc. | Doubled haploid inducer |
| WO2023076898A1 (en) | 2021-10-25 | 2023-05-04 | The Broad Institute, Inc. | Methods and compositions for editing a genome with prime editing and a recombinase |
Citations (3)
| Publication number | Priority date | Publication date | Assignee | Title |
|---|---|---|---|---|
| WO1997009436A1 (en) * | 1995-09-05 | 1997-03-13 | Crc For Biopharmaceutical Research Pty. Ltd. | Method for producing phage display vectors |
| WO1997009439A1 (en) * | 1995-09-01 | 1997-03-13 | Genvec, Inc. | Methods and vectors for site-specific recombination |
| WO1997047758A1 (en) * | 1996-06-14 | 1997-12-18 | Massachusetts Institute Of Technology | Methods and compositions for transforming cells |
Family Cites Families (52)
| Publication number | Priority date | Publication date | Assignee | Title |
|---|---|---|---|---|
| US2003A (en) * | 1841-03-12 | Improvement in horizontal windivhlls | ||
| CA1341290C (en) | 1985-05-13 | 2001-09-11 | Thomas Hohn | Method of genetically modifying plants |
| US5013659A (en) | 1987-07-27 | 1991-05-07 | E. I. Du Pont De Nemours And Company | Nucleic acid fragment encoding herbicide resistant plant acetolactate synthase |
| ATE110778T1 (en) | 1990-05-25 | 1994-09-15 | Du Pont | NUCLEOTIDE SEQUENCE OF THE GENE FOR SOYA STEAROYL-ACP DESATURASE. |
| WO1992015694A1 (en) | 1991-03-08 | 1992-09-17 | The Salk Institute For Biological Studies | Flp-mediated gene modification in mammalian cells, and compositions and cells useful therefor |
| JPH06506603A (en) | 1991-03-27 | 1994-07-28 | リサーチ コーポレーション テクノロジーズ インコーポレーテッド | single stranded circular oligonucleotide |
| WO1993001283A1 (en) | 1991-07-08 | 1993-01-21 | The United States Of America As Represented By The Secretary Of Agriculture | Selection-gene-free transgenic plants |
| DE69330227T2 (en) | 1992-02-26 | 2001-10-25 | Zeneca Mogen B V | AGROBACTERIAL TRIBES CAPABLE OF LOCATION-SPECIFIC RECOMINATION |
| DK0672142T3 (en) | 1992-12-04 | 2001-06-18 | Medical Res Council | Multivalent and multi-specific binding proteins as well as their preparation and use |
| US5527695A (en) * | 1993-01-29 | 1996-06-18 | Purdue Research Foundation | Controlled modification of eukaryotic genomes |
| US5866755A (en) * | 1993-06-14 | 1999-02-02 | Basf Aktiengellschaft | Animals transgenic for a tetracycline-regulated transcriptional inhibitor |
| EP0632054A1 (en) | 1993-06-28 | 1995-01-04 | European Molecular Biology Laboratory | Regulation of site-specific recombination by site-specific recombinase/nuclear receptor fusion proteins |
| AU690171B2 (en) | 1993-12-03 | 1998-04-23 | Medical Research Council | Recombinant binding proteins and peptides |
| NZ278490A (en) * | 1993-12-09 | 1998-03-25 | Univ Jefferson | Chimeric polynucleotide with both ribo- and deoxyribonucleotides in one strand and deoxyribonucleotides in a second strand |
| EG23907A (en) | 1994-08-01 | 2007-12-30 | Delta & Pine Land Co | Control of plant gene expression |
| US5723765A (en) * | 1994-08-01 | 1998-03-03 | Delta And Pine Land Co. | Control of plant gene expression |
| EP0937098B1 (en) | 1995-06-07 | 2002-08-14 | Invitrogen Corporation | Recombinational cloning in vitro using engineered recombination sites |
| FR2735332B1 (en) * | 1995-06-13 | 1997-07-18 | Comasec International | PERSONAL PROTECTION EQUIPMENT OF A SUPERIOR MEMBER AGAINST MECHANICAL RISKS, DEVICE AND METHOD FOR DETERMINING THE PERFORMANCE OF THIS EQUIPMENT |
| FR2736926B1 (en) | 1995-07-19 | 1997-08-22 | Rhone Poulenc Agrochimie | 5-ENOL PYRUVYLSHIKIMATE-3-PHOSPHATE SYNTHASE MUTEE, CODING GENE FOR THIS PROTEIN AND PROCESSED PLANTS CONTAINING THIS GENE |
| US6051409A (en) * | 1995-09-25 | 2000-04-18 | Novartis Finance Corporation | Method for achieving integration of exogenous DNA delivered by non-biological means to plant cells |
| CA2234557C (en) | 1995-10-13 | 2007-01-09 | Purdue Research Foundation | Method for the production of hybrid plants |
| AUPN903196A0 (en) | 1996-03-29 | 1996-04-26 | Australian National University, The | Single-step excision means |
| US5731181A (en) | 1996-06-17 | 1998-03-24 | Thomas Jefferson University | Chimeric mutational vectors having non-natural nucleotides |
| GB9711015D0 (en) | 1997-05-28 | 1997-07-23 | Zeneca Ltd | Improvements in or relating to organic compounds |
| US6774279B2 (en) * | 1997-05-30 | 2004-08-10 | Carnegie Institution Of Washington | Use of FLP recombinase in mice |
| CA2298886A1 (en) | 1997-08-05 | 1999-02-18 | Kimeragen, Inc. | The use of mixed duplex oligonucleotides to effect localized genetic changes in plants |
| US7135608B1 (en) * | 1997-08-28 | 2006-11-14 | The Salk Institute For Biological Studies | Site-specific recombination in eukaryotes and constructs useful therefor |
| US6161400A (en) * | 1997-09-23 | 2000-12-19 | Whizard Protective Wear Corp. | Cut-resistant knitted fabric |
| US6473425B1 (en) * | 1997-10-02 | 2002-10-29 | Sun Microsystems, Inc. | Mechanism for dispatching packets via a telecommunications network |
| US6114600A (en) * | 1997-10-31 | 2000-09-05 | The United States Of America As Represented By The Secretary Of Agriculture | Resolution of complex integration patterns to obtain single copy transgenes |
| WO1999025855A1 (en) | 1997-11-18 | 1999-05-27 | Pioneer Hi-Bred International, Inc. | Mobilization of viral genomes from t-dna using site-specific recombination systems |
| US7102055B1 (en) * | 1997-11-18 | 2006-09-05 | Pioneer Hi-Bred International, Inc. | Compositions and methods for the targeted insertion of a nucleotide sequence of interest into the genome of a plant |
| WO1999025840A1 (en) * | 1997-11-18 | 1999-05-27 | Pioneer Hi-Bred International, Inc. | A novel method for the integration of foreign dna into eukaryoticgenomes |
| DE69831265T2 (en) * | 1997-11-18 | 2006-06-08 | Pioneer Hi-Bred International, Inc. | COMPOSITIONS AND METHODS FOR THE GENETIC MODIFICATION OF PLANTS |
| WO1999025853A1 (en) | 1997-11-18 | 1999-05-27 | Pioneer Hi-Bred International, Inc. | Targeted manipulation of herbicide-resistance genes in plants |
| WO1999055851A2 (en) | 1998-04-28 | 1999-11-04 | Novartis Ag | Site-directed transformation of plants |
| US6781032B1 (en) * | 1998-08-14 | 2004-08-24 | The Regents Of The University Of California | Homologous recombination in plants |
| AU2982300A (en) * | 1999-02-03 | 2000-08-25 | Children's Medical Center Corporation | Gene repair involving (in vivo) excision of targeting dna |
| US6746870B1 (en) * | 1999-07-23 | 2004-06-08 | The Regents Of The University Of California | DNA recombination in eukaryotic cells by the bacteriophage PHIC31 recombination system |
| WO2001011058A1 (en) | 1999-08-09 | 2001-02-15 | Monsanto Technology Llc | Novel cloning methods and vectors |
| WO2001023545A1 (en) * | 1999-09-30 | 2001-04-05 | Sumitomo Pharmaceuticals Company, Limited | Dna containing variant frt sequences |
| US7126041B1 (en) * | 1999-12-10 | 2006-10-24 | North Carolina State Unversity | High efficiency gene targeting in plants |
| CA2416701A1 (en) * | 2000-07-21 | 2002-01-31 | The United States Of America, As Represented By The Secretary Of Agricul Ture | Methods for the replacement, translocation and stacking of dna in eukaryotic genomes |
| DE10131786A1 (en) * | 2001-07-04 | 2003-01-16 | Sungene Gmbh & Co Kgaa | Recombination systems and methods for removing nucleic acid sequences from the genome of eukaryotic organisms |
| JP4436130B2 (en) * | 2001-09-14 | 2010-03-24 | セレクティス | Random incorporation of polynucleotides by in vitro linearization |
| EP1572914B1 (en) * | 2002-03-29 | 2012-06-06 | Syngenta Participations AG | Lambda integrase mediated recombination in plants |
| ATE419367T1 (en) * | 2003-11-18 | 2009-01-15 | Bayer Bioscience Nv | IMPROVED TARGETED DNA INSERTION IN PLANTS |
| US7935862B2 (en) * | 2003-12-02 | 2011-05-03 | Syngenta Participations Ag | Targeted integration and stacking of DNA through homologous recombination |
| US20060288444A1 (en) * | 2004-08-13 | 2006-12-21 | Mccarroll Robert | Soybean polymorphisms and methods of genotyping |
| PT1885176T (en) * | 2005-05-27 | 2016-11-28 | Monsanto Technology Llc | Soybean event mon89788 and methods for detection thereof |
| US20080083042A1 (en) * | 2006-08-14 | 2008-04-03 | David Butruille | Maize polymorphisms and methods of genotyping |
| AR067114A1 (en) * | 2007-06-22 | 2009-09-30 | Monsanto Technology Llc | METHODS AND COMPOSITIONS TO SELECT LOCI BASED ON PERFORMANCE AND EXCESSION OF CHARACTERISTICS |
-
1998
- 1998-11-17 DE DE69831265T patent/DE69831265T2/en not_active Expired - Lifetime
- 1998-11-17 AT AT05006893T patent/ATE401410T1/en not_active IP Right Cessation
- 1998-11-17 AT AT98958629T patent/ATE302271T1/en not_active IP Right Cessation
- 1998-11-17 EP EP98958629A patent/EP1034262B1/en not_active Expired - Lifetime
- 1998-11-17 NZ NZ503859A patent/NZ503859A/en unknown
- 1998-11-17 EP EP98960262A patent/EP1032693A1/en not_active Withdrawn
- 1998-11-17 CA CA002305866A patent/CA2305866A1/en not_active Abandoned
- 1998-11-17 AU AU14629/99A patent/AU760113C/en not_active Ceased
- 1998-11-17 ES ES98958629T patent/ES2245487T3/en not_active Expired - Lifetime
- 1998-11-17 US US09/193,502 patent/US6187994B1/en not_active Expired - Lifetime
- 1998-11-17 NZ NZ504300A patent/NZ504300A/en unknown
- 1998-11-17 US US09/193,484 patent/US6331661B1/en not_active Expired - Lifetime
- 1998-11-17 PT PT98958629T patent/PT1034262E/en unknown
- 1998-11-17 EP EP05006893A patent/EP1574573B9/en not_active Expired - Lifetime
- 1998-11-17 CA CA002306184A patent/CA2306184C/en not_active Expired - Fee Related
- 1998-11-17 AU AU15905/99A patent/AU757672B2/en not_active Ceased
- 1998-11-17 WO PCT/US1998/024609 patent/WO1999025854A1/en not_active Application Discontinuation
- 1998-11-17 DE DE69839742T patent/DE69839742D1/en not_active Expired - Lifetime
- 1998-11-17 WO PCT/US1998/024610 patent/WO1999025821A1/en active IP Right Grant
- 1998-11-17 ES ES05006893T patent/ES2308327T3/en not_active Expired - Lifetime
- 1998-11-17 DK DK98958629T patent/DK1034262T3/en active
-
1999
- 1999-11-12 US US09/439,042 patent/US6573425B1/en not_active Expired - Lifetime
- 1999-11-12 US US09/438,874 patent/US6455315B1/en not_active Expired - Lifetime
- 1999-11-12 US US09/439,158 patent/US6458594B1/en not_active Expired - Lifetime
- 1999-11-12 US US09/438,239 patent/US6552248B1/en not_active Expired - Lifetime
- 1999-12-06 US US09/455,050 patent/US6624297B1/en not_active Expired - Lifetime
-
2003
- 2003-03-24 AU AU2003202440A patent/AU2003202440B8/en not_active Ceased
- 2003-05-07 US US10/430,908 patent/US7405079B2/en not_active Expired - Fee Related
- 2003-05-07 US US10/430,907 patent/US7572634B2/en not_active Expired - Fee Related
- 2003-05-16 US US10/440,030 patent/US7361508B2/en not_active Expired - Fee Related
- 2003-08-12 US US10/639,751 patent/US9222098B2/en not_active Expired - Fee Related
-
2008
- 2008-02-28 US US12/039,307 patent/US8735158B2/en not_active Expired - Fee Related
- 2008-11-06 US US12/266,016 patent/US7820880B2/en not_active Expired - Fee Related
Patent Citations (3)
| Publication number | Priority date | Publication date | Assignee | Title |
|---|---|---|---|---|
| WO1997009439A1 (en) * | 1995-09-01 | 1997-03-13 | Genvec, Inc. | Methods and vectors for site-specific recombination |
| WO1997009436A1 (en) * | 1995-09-05 | 1997-03-13 | Crc For Biopharmaceutical Research Pty. Ltd. | Method for producing phage display vectors |
| WO1997047758A1 (en) * | 1996-06-14 | 1997-12-18 | Massachusetts Institute Of Technology | Methods and compositions for transforming cells |
Non-Patent Citations (3)
| Title |
|---|
| B. SAUER: "Identification of cryptic lox sites in the yeast genome by selection for Cre-mediated chromosome translocations that confer multiple drug resistance", JOURNAL OF MOLECULAR BIOLOGY, vol. 223, 1992, pages 911 - 928, XP002097373 * |
| J. SEIBLER ET AL.: "Double reciprocal crossover by FLP-recombinase: a concept and an assay", BIOCHEMISTRY, vol. 36, 1997, pages 1740 - 1747, XP002074110 * |
| J.F. SENECOFF ET AL.: "DNA recognition by the FLP recombinase of the yeast 2 micron plasmid", JOURNAL OF MOLECULAR BIOLOGY, vol. 201, 1988, pages 405 - 421, XP002097374 * |
Cited By (313)
| Publication number | Priority date | Publication date | Assignee | Title |
|---|---|---|---|---|
| WO1999055851A3 (en) * | 1998-04-28 | 2000-01-20 | Novartis Ag | Site-directed transformation of plants |
| WO1999055851A2 (en) * | 1998-04-28 | 1999-11-04 | Novartis Ag | Site-directed transformation of plants |
| US6746870B1 (en) | 1999-07-23 | 2004-06-08 | The Regents Of The University Of California | DNA recombination in eukaryotic cells by the bacteriophage PHIC31 recombination system |
| US8129598B2 (en) | 1999-07-23 | 2012-03-06 | The Regents Of The University Of California | DNA recombination in eukaryotic cells by the bacteriophage PHIC31 recombination system |
| EP2161340A2 (en) | 1999-12-16 | 2010-03-10 | CropDesign N.V. | Optimized T-DNA transfer and vectors therefor |
| WO2001085969A3 (en) * | 2000-05-08 | 2002-07-04 | Pioneer Hi Bred Int | Genetic transformation in plants using site-specific recombination and wide hybridization |
| WO2001085969A2 (en) * | 2000-05-08 | 2001-11-15 | Pioneer Hi-Bred International, Inc. | Genetic transformation in plants using site-specific recombination and wide hybridization |
| WO2002006500A2 (en) * | 2000-07-18 | 2002-01-24 | Pioneer Hi-Bred International, Inc. | Methods of transforming plants and identifying parental origin of a chromosome in those plants |
| WO2002006500A3 (en) * | 2000-07-18 | 2002-12-27 | Pioneer Hi Bred Int | Methods of transforming plants and identifying parental origin of a chromosome in those plants |
| US6936747B2 (en) | 2000-07-21 | 2005-08-30 | The United States Of America As Represented By The Secretary Of Agriculture | Methods for the replacement, translocation and stacking of DNA in eukaryotic genomes |
| WO2002008409A2 (en) * | 2000-07-21 | 2002-01-31 | The United States Of America, As Represented By The Secretary Of Agriculture | Methods for the replacement, translocation and stacking of dna in eukaryotic genomes |
| US7972857B2 (en) | 2000-07-21 | 2011-07-05 | The United States Of America As Represented By The Secretary Of Agriculture | Methods for the replacement, translocation and stacking of DNA in eukaryotic genomes |
| WO2002008409A3 (en) * | 2000-07-21 | 2003-03-13 | Us Agriculture | Methods for the replacement, translocation and stacking of dna in eukaryotic genomes |
| EP2039769A2 (en) | 2000-09-13 | 2009-03-25 | Pioneer Hi-Bred International, Inc. | Antimicrobial peptides and methods of use |
| US7763458B2 (en) | 2000-10-06 | 2010-07-27 | Icon Genetics Gmbh | Vector system for plants |
| US7652194B2 (en) | 2000-12-08 | 2010-01-26 | Icon Genetics Gmbh | Processes and vectors for producing transgenic plants |
| US7193131B2 (en) | 2001-01-19 | 2007-03-20 | Icon Genetics Ag | Processes and vectors for plastid transformation of higher plants |
| US8058506B2 (en) | 2001-03-23 | 2011-11-15 | Icon Genetics Gmbh | Site-targeted transformation using amplification vectors |
| WO2002077246A3 (en) * | 2001-03-23 | 2003-11-20 | Icon Genetics Ag | Site-targeted transformation using amplification vectors |
| US7667091B2 (en) | 2001-03-29 | 2010-02-23 | Icon Genetics Gmbh | Method of encoding information in nucleic acids of a genetically engineered organism |
| US7667092B2 (en) | 2001-04-30 | 2010-02-23 | Icon Genetics Gmbh | Processes and vectors for amplification or expression of nucleic acid sequences of interest in plants |
| US7521240B2 (en) | 2001-05-30 | 2009-04-21 | Smithkline Beecham Corporation | Chromosome-based platforms |
| US7371923B2 (en) | 2001-07-06 | 2008-05-13 | Icon Genetics Ag | Process of generating transplastomic plants or plant cells devoid of a selection marker |
| US8257945B2 (en) | 2001-09-04 | 2012-09-04 | Icon Genetics, Inc. | Identification of eukaryotic internal ribosome entry site (IRES) elements |
| US8192984B2 (en) | 2001-09-04 | 2012-06-05 | Icon Genetics, Inc. | Creation of artificial internal ribosome entry site (IRES) elements |
| EP1961293A2 (en) | 2001-09-27 | 2008-08-27 | Pioneer Hi-Bred International, Inc. | Phytate polynucleotides and methods of use |
| US7351877B2 (en) | 2002-03-29 | 2008-04-01 | Syngenta Participations Ag | Lambda integrase mediated recombination in plants |
| WO2003087313A2 (en) | 2002-04-08 | 2003-10-23 | Pioneer Hi-Bred International, Inc. | Enhanced silk exsertion under stress |
| EP1499720A2 (en) * | 2002-05-03 | 2005-01-26 | Pioneer Hi-Bred International, Inc. | Gene targeting using replicating dna molecules |
| AU2003234330B2 (en) * | 2002-05-03 | 2008-06-19 | Pioneer Hi-Bred International, Inc. | Gene targeting using replicating DNA molecules |
| EP1499720A4 (en) * | 2002-05-03 | 2005-11-09 | Pioneer Hi Bred Int | Gene targeting using replicating dna molecules |
| US7164056B2 (en) | 2002-05-03 | 2007-01-16 | Pioneer Hi-Bred International, Inc. | Gene targeting using replicating DNA molecules |
| US7608752B2 (en) | 2002-05-03 | 2009-10-27 | Pioneer Hi-Bred International, Inc. | Gene targeting using replicating DNA molecules |
| EP2275536A1 (en) | 2002-08-06 | 2011-01-19 | Verdia, Inc. | AP1 amine oxidase variants |
| EP2322629A2 (en) | 2003-04-29 | 2011-05-18 | Pioneer Hi-Bred International Inc. | Novel glyphosate-n-acetyltransferase (GAT) genes |
| EP2535414A1 (en) | 2003-04-29 | 2012-12-19 | Pioneer Hi-Bred International Inc. | Novel glyphosate-n-acetyltransferase (gat) genes |
| WO2005016504A2 (en) | 2003-06-23 | 2005-02-24 | Pioneer Hi-Bred International, Inc. | Disruption of acc synthase genes to delay senescence in plants |
| EP2361984A1 (en) | 2003-10-09 | 2011-08-31 | E. I. du Pont de Nemours and Company | Gene silencing by using modified micro-RNA molecules |
| WO2005035769A2 (en) | 2003-10-09 | 2005-04-21 | E. I. Du Pont De Nemours And Company | Gene silencing by using micro-rna molecules |
| US7935862B2 (en) | 2003-12-02 | 2011-05-03 | Syngenta Participations Ag | Targeted integration and stacking of DNA through homologous recombination |
| US8399254B2 (en) | 2003-12-02 | 2013-03-19 | Syngenta Participations Ag | Targeted integration of DNA through recombination |
| WO2005059148A1 (en) * | 2003-12-17 | 2005-06-30 | Pioneer Hi-Bred International, Inc. | Recombinase mediated gene traps |
| EP2298915A1 (en) | 2004-06-30 | 2011-03-23 | Pioneer Hi-Bred International, Inc. | Methods of protecting plants from pathogenic fungi |
| EP2298914A1 (en) | 2004-06-30 | 2011-03-23 | Pioneer Hi-Bred International, Inc. | Methods of protecting plants from pathogenic fungi |
| EP2264053A2 (en) | 2004-07-02 | 2010-12-22 | Pioneer-Hi-Bred International, Inc. | Antifungal polypeptides |
| EP2113512A2 (en) | 2004-07-02 | 2009-11-04 | Pioneer Hi-Bred International Inc. | Antifungal polypeptides |
| EP2230247A2 (en) | 2004-07-02 | 2010-09-22 | Pioneer-Hi-Bred International, Inc. | Antifungal polypeptides |
| EP2003205A2 (en) | 2004-12-28 | 2008-12-17 | Pioneer Hi-Bred International, Inc. | Improved grain quality through altered expression of seed proteins |
| EP2338905A2 (en) | 2005-02-23 | 2011-06-29 | North Carolina State University | Alteration of tobacco alkaloid content through modification of specific cytochrome p450 genes |
| EP2261362A2 (en) | 2005-05-25 | 2010-12-15 | Pioneer Hi-Bred International Inc. | Methods for improving crop plant architecture and yield |
| EP2261361A2 (en) | 2005-05-25 | 2010-12-15 | Pioneer Hi-Bred International Inc. | Methods for improving crop plant architecture and yield |
| WO2007011733A3 (en) * | 2005-07-18 | 2007-09-07 | Pioneer Hi Bred Int | Modified frt recombination sites and methods of use |
| US9234194B2 (en) | 2005-07-18 | 2016-01-12 | Pioneer Hi-Bred International, Inc. | Modified FRT recombination site libraries and methods of use |
| US8586361B2 (en) | 2005-07-18 | 2013-11-19 | Pioneer Hi-Bred International, Inc. | FRT recombination sites and methods of use |
| US8318493B2 (en) | 2005-07-18 | 2012-11-27 | Pioneer Hi-Bred International, Inc. | FRT recombination sites and methods of use |
| US7736897B2 (en) | 2005-07-18 | 2010-06-15 | Pioneer Hi-Bred International, Inc. | FRT recombination sites and methods of use |
| WO2007011733A2 (en) | 2005-07-18 | 2007-01-25 | Pioneer Hi-Bred International, Inc. | Modified frt recombination sites and methods of use |
| US8900869B2 (en) | 2005-07-18 | 2014-12-02 | Pioneer Hi-Bred International, Inc. | FRT recombination sites and methods of use |
| US9777284B2 (en) | 2005-07-18 | 2017-10-03 | Pioneer Hi-Bred International, Inc. | Modified FRT recombination site libraries and methods of use |
| EP2112223A2 (en) | 2005-11-10 | 2009-10-28 | Pioneer Hi-Bred International Inc. | DOF (DNA binding with one finger) sequences and method of use |
| WO2007059077A2 (en) | 2005-11-14 | 2007-05-24 | E.I.Du Pont De Nemours And Company | Compositions and methods for altering alpha- and beta-tocotrienol content |
| WO2007079161A2 (en) | 2005-12-30 | 2007-07-12 | Pioneer Hi-Bred International, Inc. | Udp-xylose synthases (uxs) polynucleotides, polypeptides, and uses thereof |
| EP2730587A2 (en) | 2006-02-09 | 2014-05-14 | Pioneer Hi-Bred International, Inc. | Genes for enhancing nitrogen utilization efficiency in crop plants |
| WO2007103738A2 (en) | 2006-03-01 | 2007-09-13 | Pioneer Hi-Bred International, Inc. | Compositions related to the quantitative trait locus 6 (qtl6) in maize and methods of use |
| EP2251349A1 (en) | 2006-04-19 | 2010-11-17 | Pioneer Hi-Bred International, Inc. | Isolated polynucleotide molecules corresponding to mutant and wild-type alleles of the maize D9 gene and methods of use |
| EP2333088A1 (en) | 2006-05-16 | 2011-06-15 | Pioneer Hi-Bred International, Inc. | Antifungal polypeptides |
| EP2308986A1 (en) | 2006-05-17 | 2011-04-13 | Pioneer Hi-Bred International Inc. | Artificial plant minichromosomes |
| EP2380987A2 (en) | 2006-06-28 | 2011-10-26 | Pioneer Hi-Bred International Inc. | Soybean event 3560.4.3.5 and compositions and methods for the identification and/or detection thereof |
| WO2008002872A2 (en) | 2006-06-28 | 2008-01-03 | Pioneer Hi-Bred International, Inc. | Soybean event 3560.4.3.5 and compositions and methods for the identification and/or detection thereof |
| EP2664673A2 (en) | 2006-10-05 | 2013-11-20 | E. I. du Pont de Nemours and Company | Maize microRNA sequences |
| US9290777B2 (en) | 2007-02-05 | 2016-03-22 | National University Of Singapore | Putative cytokinin receptor and methods for use thereof |
| US8445746B2 (en) | 2007-02-23 | 2013-05-21 | University Of Georgia Research Foundation, Inc. | Compositions and methods for identifying genetic sequences with toxin resistance in plants |
| WO2008112970A2 (en) | 2007-03-14 | 2008-09-18 | Pioneer Hi-Bred International, Inc. | Dominant gene suppression transgenes and methods of using same |
| WO2008145629A2 (en) | 2007-05-25 | 2008-12-04 | Cropdesign N.V. | Yield enhancement in plants by modulation of maize alfins |
| EP2568048A1 (en) | 2007-06-29 | 2013-03-13 | Pioneer Hi-Bred International, Inc. | Methods for altering the genome of a monocot plant cell |
| WO2009067580A2 (en) | 2007-11-20 | 2009-05-28 | Pioneer Hi-Bred International, Inc. | Maize ethylene signaling genes and modulation of same for improved stress tolerance in plants |
| EP2405013A2 (en) | 2008-01-17 | 2012-01-11 | Pioneer Hi-Bred International Inc. | Compositions and methods for the suppression of target polynucleotides from the family Aphididae |
| EP2548964A2 (en) | 2008-01-17 | 2013-01-23 | Pioneer Hi-Bred International Inc. | Compositions and methods for the suppression of target polynucleotides from Lepidoptera |
| US7964774B2 (en) | 2008-05-14 | 2011-06-21 | Monsanto Technology Llc | Plants and seeds of spring canola variety SCV384196 |
| EP3441470A1 (en) | 2008-09-26 | 2019-02-13 | BASF Agrochemical Products, B.V. | Herbicide-resistant ahas-mutants and methods of use |
| EP2821490A2 (en) | 2008-10-30 | 2015-01-07 | Pioneer Hi-Bred International Inc. | Manipulation of glutamine synthetases (GS) to improve nitrogen use efficiency and grain yield in higher plants |
| WO2010065867A1 (en) | 2008-12-04 | 2010-06-10 | Pioneer Hi-Bred International, Inc. | Methods and compositions for enhanced yield by targeted expression of knotted1 |
| US8574910B2 (en) | 2008-12-19 | 2013-11-05 | E. I. Du Pont De Nemours And Company | Site-specific integration and stacking of transgenes in soybean via DNA recombinase mediated cassette exchange |
| WO2010080430A1 (en) * | 2008-12-19 | 2010-07-15 | E. I. Du Pont De Nemours And Company | Site-specific integration and stacking of transgenes in soybean via dna recombinase mediated cassette exchange |
| US8293533B2 (en) | 2008-12-19 | 2012-10-23 | E.I. Du Pont De Nemours And Company | Site-specific integration and stacking of transgenes in soybean via DNA recombinase mediated cassette exchange |
| WO2010078445A1 (en) | 2008-12-31 | 2010-07-08 | Pioneer Hi-Bred International, Inc. | Auxotrophic agrobacterium for plant transformation and methods thereof |
| WO2010083328A2 (en) | 2009-01-14 | 2010-07-22 | The Salk Institute For Biological Studies | Methods for screening and compounds that protect against amyloid diseases |
| EP3156489A1 (en) | 2009-01-22 | 2017-04-19 | Syngenta Participations Ag | Mutant hydroxyphenylpyruvate dioxygenase polypeptides and methods of use |
| EP2573183A1 (en) | 2009-01-22 | 2013-03-27 | Syngenta Participations AG. | Mutant hydroxyphenylpyruvate dioxgenase polypeptids and methods of use |
| WO2010096613A1 (en) | 2009-02-19 | 2010-08-26 | Pioneer Hi-Bred International, Inc. | Blended refuge deployment via manipulation during hybrid seed production |
| WO2010101818A1 (en) | 2009-03-02 | 2010-09-10 | Pioneer Hi-Bred International, Inc. | Nac transcriptional activators involved in abiotic stress tolerance |
| WO2010120862A1 (en) | 2009-04-14 | 2010-10-21 | Pioneer Hi-Bred International, Inc. | Modulation of acc synthase improves plant yield under low nitrogen conditions |
| EP2666781A1 (en) | 2009-05-04 | 2013-11-27 | Pioneer Hi-Bred International, Inc. | Yield enhancement in plants by modulation of AP2 transcription factor |
| WO2010147825A1 (en) | 2009-06-09 | 2010-12-23 | Pioneer Hi-Bred International, Inc. | Early endosperm promoter and methods of use |
| WO2011068567A1 (en) | 2009-07-10 | 2011-06-09 | Syngenta Participations Ag | Novel hydroxyphenylpyruvate dioxygenase polypeptides and methods of use |
| WO2011011273A1 (en) | 2009-07-24 | 2011-01-27 | Pioneer Hi-Bred International, Inc. | The use of dimerization domain component stacks to modulate plant architecture |
| WO2011017492A2 (en) | 2009-08-05 | 2011-02-10 | Pioneer Hi-Bred International, Inc. | Novel eto1 genes and use of same for reduced ethylene and improved stress tolerance in plants |
| WO2011022608A2 (en) | 2009-08-20 | 2011-02-24 | Pioneer Hi-Bred International, Inc. | Functional expression of yeast nitrate transporter (ynt1) in maize to improve nitrate uptake |
| WO2011022597A1 (en) | 2009-08-20 | 2011-02-24 | Pioneer Hi-Bred International, Inc. | Functional expression of shuffled yeast nitrate transporter (ynti) in maize to improve nitrate uptake under low nitrate environment |
| WO2011021171A1 (en) | 2009-08-21 | 2011-02-24 | Beeologics, Llc | Preventing and curing beneficial insect diseases via plant transcribed molecules |
| WO2011025860A1 (en) | 2009-08-28 | 2011-03-03 | E. I. Du Pont De Nemours And Company | Compositions and methods to control insect pests |
| EP3401404A1 (en) | 2009-08-28 | 2018-11-14 | E. I. du Pont de Nemours and Company | Compositions and methods to control insect pests |
| EP3098317A1 (en) | 2009-08-28 | 2016-11-30 | E. I. du Pont de Nemours and Company | Compositions and methods to control insect pests |
| WO2011036569A2 (en) | 2009-09-22 | 2011-03-31 | The Board Of Trustees Of The University Of Arkansas, N.A. | Rice cultivar cl 142-ar |
| WO2011041796A1 (en) | 2009-10-02 | 2011-04-07 | Pioneer Hi-Bred International, Inc. | Down-regulation of acc synthase for improved plant performance |
| WO2011047191A1 (en) | 2009-10-15 | 2011-04-21 | The Board Of Trustees Of The University Of Arkansas, N.A. | Rice cultivar cl181-ar |
| WO2011056544A1 (en) | 2009-10-26 | 2011-05-12 | Pioneer Hi-Bred International, Inc. | Somatic ovule specific promoter and methods of use |
| US8704041B2 (en) | 2009-12-30 | 2014-04-22 | Pioneer Hi Bred International Inc | Methods and compositions for targeted polynucleotide modification |
| US9926571B2 (en) | 2009-12-30 | 2018-03-27 | Pioneer Hi-Bred International, Inc. | Methods and compositions for targeted polynucleotide modification |
| WO2011082318A2 (en) | 2009-12-30 | 2011-07-07 | Pioneer Hi-Bred International, Inc. | Methods and compositions for the introduction and regulated expression of genes in plants |
| WO2011082310A2 (en) | 2009-12-30 | 2011-07-07 | Pioneer Hi-Bred International, Inc. | Methods and compositions for targeted polynucleotide modification |
| US10443064B2 (en) | 2009-12-30 | 2019-10-15 | Pioneer Hi-Bred International, Inc. | Methods and compositions for targeted polynucleotide modification |
| EP3078748A2 (en) | 2009-12-30 | 2016-10-12 | Pioneer Hi-Bred International, Inc. | Methods and compositions for the introduction and regulated expression of genes in plants |
| WO2011082304A1 (en) | 2009-12-31 | 2011-07-07 | Pioneer Hi-Bred International, Inc. | Engineering plant resistance to diseases caused by pathogens |
| WO2011085062A1 (en) | 2010-01-06 | 2011-07-14 | Pioneer Hi-Bred International, Inc. | Identification of diurnal rhythms in photosynthetic and non-photosynthetic tissues from zea mays and use in improving crop plants |
| WO2011094199A1 (en) | 2010-01-26 | 2011-08-04 | Pioneer Hi-Bred International, Inc. | Polynucleotide and polypeptide sequences associated with herbicide tolerance |
| WO2011094205A1 (en) | 2010-01-26 | 2011-08-04 | Pioneer Hi-Bred International, Inc. | Hppd-inhibitor herbicide tolerance |
| WO2011139431A1 (en) | 2010-05-06 | 2011-11-10 | Pioneer Hi-Bred International, Inc. | Maize acc synthase 3 gene and protein and uses thereof |
| WO2011163590A1 (en) | 2010-06-25 | 2011-12-29 | E. I. Du Pont De Nemours And Company | Compositions and methods for enhancing resistance to northern leaf blight in maize |
| EP3366117A1 (en) | 2010-06-25 | 2018-08-29 | E. I. du Pont de Nemours and Company | Compositions and methods for enhancing resistance to northern leaf blight in maize |
| WO2012021785A1 (en) | 2010-08-13 | 2012-02-16 | Pioneer Hi-Bred International, Inc. | Compositions and methods comprising sequences having hydroxyphenylpyruvate dioxygenase (hppd) activity |
| WO2012021794A1 (en) | 2010-08-13 | 2012-02-16 | Pioneer Hi-Bred International, Inc. | Chimeric promoters and methods of use |
| WO2012021797A1 (en) | 2010-08-13 | 2012-02-16 | Pioneer Hi-Bred International, Inc. | Methods and compositions for targeting sequences of interest to the chloroplast |
| WO2012027209A2 (en) | 2010-08-23 | 2012-03-01 | Pioneer Hi-Bred International, Inc. | Novel defensin variants and methods of use |
| WO2012071040A1 (en) | 2010-11-24 | 2012-05-31 | Pioneer Hi-Bred International, Inc. | Brassica gat event dp-073496-4 and compositions and methods for the identification and/or detection thereof |
| WO2012071039A1 (en) | 2010-11-24 | 2012-05-31 | Pioner Hi-Bred International, Inc. | Brassica gat event dp-061061-7 and compositions and methods for the identification and/or detection thereof |
| WO2012078949A2 (en) | 2010-12-09 | 2012-06-14 | Syngenta Participations Ag | Methods and compositions using small interfering rna (sirna) for nematode control in plants |
| WO2012082548A2 (en) | 2010-12-15 | 2012-06-21 | Syngenta Participations Ag | Soybean event syht0h2 and compositions and methods for detection thereof |
| WO2012092106A1 (en) | 2010-12-28 | 2012-07-05 | Pioneer Hi-Bred International, Inc. | Novel bacillus thuringiensis gene with lepidopteran activity |
| WO2012106321A1 (en) | 2011-02-01 | 2012-08-09 | Colorado Wheat Research Foundation, Inc. | Acetyl co-enzyme a carboxylase herbicide resistant plants |
| EP3682731A1 (en) | 2011-02-01 | 2020-07-22 | Colorado Wheat Research Foundation, Inc. | Acetyl co-enzyme a carboxylase herbicide resistant plants |
| WO2012109515A1 (en) | 2011-02-11 | 2012-08-16 | Pioneer Hi-Bred International, Inc. | Synthetic insecticidal proteins active against corn rootworm |
| WO2012112411A1 (en) | 2011-02-15 | 2012-08-23 | Pioneer Hi-Bred International, Inc. | Root-preferred promoter and methods of use |
| WO2012122369A1 (en) | 2011-03-10 | 2012-09-13 | Pioneer Hi-Bred International, Inc. | Novel bacillus thuringiensis gene with lepidopteran activity |
| US10822610B2 (en) | 2011-03-23 | 2020-11-03 | E. I. Du Pont De Nemours And Company | Methods for producing a complex transgenic trait locus |
| WO2012129373A2 (en) | 2011-03-23 | 2012-09-27 | Pioneer Hi-Bred International, Inc. | Methods for producing a complex transgenic trait locus |
| WO2012131495A2 (en) | 2011-03-30 | 2012-10-04 | Universidad Nacional Autónoma de México | Mutant bacillus thuringiensis cry genes and methods of use |
| WO2012142311A1 (en) | 2011-04-15 | 2012-10-18 | Pioneer Hi-Bred International, Inc. | Self-reproducing hybrid plants |
| US8685737B2 (en) | 2011-04-27 | 2014-04-01 | Amyris, Inc. | Methods for genomic modification |
| US9701971B2 (en) | 2011-04-27 | 2017-07-11 | Amyris, Inc. | Methods for genomic modification |
| WO2012149470A1 (en) | 2011-04-27 | 2012-11-01 | Amyris, Inc. | Methods for genomic modification |
| WO2012148835A1 (en) | 2011-04-29 | 2012-11-01 | Pioneer Hi-Bred International, Inc. | Down-regulation of a homeodomain-leucine zipper i-class homeobox gene for improved plant performance |
| WO2012154824A1 (en) | 2011-05-09 | 2012-11-15 | E. I. Du Pont De Nemours And Company | Methods and compositions for silencing gene families using artificial micrornas |
| WO2012161982A1 (en) | 2011-05-23 | 2012-11-29 | E. I. Dupont De Nemours & Company | Chloroplast transit peptides and methods of their use |
| WO2012174139A2 (en) | 2011-06-14 | 2012-12-20 | Synthon Biopharmaceuticals B.V. | Compositions and methods for making and b ioc ont aining auxotrophic transgenic plants |
| US9574208B2 (en) | 2011-06-21 | 2017-02-21 | Ei Du Pont De Nemours And Company | Methods and compositions for producing male sterile plants |
| WO2013066423A2 (en) | 2011-06-21 | 2013-05-10 | Pioneer Hi-Bred International, Inc. | Methods and compositions for producing male sterile plants |
| WO2013019411A1 (en) | 2011-08-03 | 2013-02-07 | E. I. Du Pont De Nemours And Company | Methods and compositions for targeted integration in a plant |
| US8785729B2 (en) | 2011-08-09 | 2014-07-22 | Nunhems, B.V. | Lettuce variety redglace |
| WO2013033308A2 (en) | 2011-08-31 | 2013-03-07 | Pioneer Hi-Bred International, Inc. | Methods for tissue culture and transformation of sugarcane |
| US8754293B2 (en) | 2011-09-09 | 2014-06-17 | Nunhems B.V. | Lettuce variety intred |
| WO2013063344A1 (en) | 2011-10-28 | 2013-05-02 | Pioneer Hi-Bred International, Inc. | Engineered pep carboxylase variants for improved plant productivity |
| WO2013063487A1 (en) | 2011-10-28 | 2013-05-02 | E. I. Du Pont De Nemours And Company | Methods and compositions for silencing genes using artificial micrornas |
| WO2013066805A1 (en) | 2011-10-31 | 2013-05-10 | Pioneer Hi-Bred International, Inc. | Improving plant drought tolerance, nitrogen use efficiency and yield |
| WO2013096810A1 (en) | 2011-12-21 | 2013-06-27 | The Curators Of The University Of Missouri | Soybean variety s05-11482 |
| WO2013096818A1 (en) | 2011-12-21 | 2013-06-27 | The Curators Of The University Of Missouri | Soybean variety s05-11268 |
| US9380756B2 (en) | 2012-01-04 | 2016-07-05 | Nunhems B.V. | Lettuce variety multigreen 50 |
| WO2013103366A1 (en) | 2012-01-06 | 2013-07-11 | Pioneer Hi-Bred International, Inc. | A method to screen plants for genetic elements inducing parthenogenesis in plants |
| WO2013103371A1 (en) | 2012-01-06 | 2013-07-11 | Pioneer Hi-Bred International, Inc. | Ovule specific promoter and methods of use |
| WO2013103365A1 (en) | 2012-01-06 | 2013-07-11 | Pioneer Hi-Bred International, Inc. | Pollen preferred promoters and methods of use |
| WO2013104026A1 (en) | 2012-01-11 | 2013-07-18 | The Australian National University | Method for modulating plant root architecture |
| WO2013112686A1 (en) | 2012-01-27 | 2013-08-01 | Pioneer Hi-Bred International, Inc. | Methods and compositions for generating complex trait loci |
| US10435699B2 (en) | 2012-01-27 | 2019-10-08 | E I Du Pont De Nemours And Company | Methods and compositions for generating complex trait loci |
| WO2013122720A2 (en) | 2012-02-16 | 2013-08-22 | Syngenta Participations Ag | Engineered pesticidal proteins |
| WO2013138358A1 (en) | 2012-03-13 | 2013-09-19 | Pioneer Hi-Bred International, Inc. | Genetic reduction of male fertility in plants |
| WO2013138309A1 (en) | 2012-03-13 | 2013-09-19 | Pioneer Hi-Bred International, Inc. | Genetic reduction of male fertility in plants |
| US8835720B2 (en) | 2012-04-26 | 2014-09-16 | Monsanto Technology Llc | Plants and seeds of spring canola variety SCV967592 |
| US8878009B2 (en) | 2012-04-26 | 2014-11-04 | Monsanto Technology, LLP | Plants and seeds of spring canola variety SCV318181 |
| US8859857B2 (en) | 2012-04-26 | 2014-10-14 | Monsanto Technology Llc | Plants and seeds of spring canola variety SCV259778 |
| WO2013166113A1 (en) | 2012-05-04 | 2013-11-07 | E. I. Du Pont De Nemours And Company | Compositions and methods comprising sequences having meganuclease activity |
| WO2013177400A1 (en) | 2012-05-24 | 2013-11-28 | Iowa State University Research Foundation, Inc. | Arabidopsis nonhost resistance gene(s) and use thereof to engineer disease resistant plants |
| WO2013188291A2 (en) | 2012-06-15 | 2013-12-19 | E. I. Du Pont De Nemours And Company | Methods and compositions involving als variants with native substrate preference |
| WO2013188501A1 (en) | 2012-06-15 | 2013-12-19 | Pioneer Hi-Bred International, Inc. | Genetic loci associated with resistance of soybean to cyst nematode and methods of use |
| WO2013192256A1 (en) | 2012-06-22 | 2013-12-27 | Syngenta Participations Ag | Biological control of coleopteran pests |
| US9816102B2 (en) | 2012-09-13 | 2017-11-14 | Indiana University Research And Technology Corporation | Compositions and systems for conferring disease resistance in plants and methods of use thereof |
| WO2014059155A1 (en) | 2012-10-11 | 2014-04-17 | Pioneer Hi-Bred International, Inc. | Guard cell promoters and uses thereof |
| WO2014062544A2 (en) | 2012-10-15 | 2014-04-24 | Pioneer Hi-Bred International, Inc. | Methods and compositions to enhance activity of cry endotoxins |
| WO2014081673A2 (en) | 2012-11-20 | 2014-05-30 | Pioneer Hi-Bred International, Inc. | Engineering plants for efficient uptake and utilization of urea to improve crop production |
| WO2014143304A1 (en) | 2012-12-13 | 2014-09-18 | Pioneer Hi-Bred International, Inc. | Methods and compositions for producing and selecting transgenic plants |
| WO2014093485A1 (en) | 2012-12-13 | 2014-06-19 | Pioneer Hi-Bred International, Inc. | Methods and compositions for producing and selecting transgenic plants |
| WO2014100525A2 (en) | 2012-12-21 | 2014-06-26 | Pioneer Hi-Bred International, Inc. | Compositions and methods for auxin-analog conjugation |
| WO2014116989A1 (en) | 2013-01-25 | 2014-07-31 | Pioneer Hi-Bred International, Inc. | Maize event dp-032218-9 and methods for detection thereof |
| WO2014164775A1 (en) | 2013-03-11 | 2014-10-09 | Pioneer Hi-Bred International, Inc. | Methods and compositions to improve the spread of chemical signals in plants |
| WO2014164014A1 (en) | 2013-03-11 | 2014-10-09 | Pioneer Hi-Bred International, Inc. | Genes for improving nutrient uptake and abiotic stress tolerance in plants |
| WO2014164828A2 (en) | 2013-03-11 | 2014-10-09 | Pioneer Hi-Bred International, Inc. | Methods and compositions employing a sulfonylurea-dependent stabilization domain |
| WO2014164961A2 (en) | 2013-03-12 | 2014-10-09 | Pioneer Hi-Bred International, Inc. | Manipulation of dominant male sterility |
| WO2014159113A1 (en) | 2013-03-12 | 2014-10-02 | Pioneer Hi-Bred International, Inc. | Root-preferred promoter and methods of use |
| WO2014164466A1 (en) | 2013-03-12 | 2014-10-09 | E. I. Du Pont De Nemours And Company | Methods for the identification of variant recognition sites for rare-cutting engineered double-strand-break-inducing agents and compositions and uses thereof |
| WO2014164399A1 (en) | 2013-03-12 | 2014-10-09 | Pioneer Hi-Bred International, Inc. | Root-preferred promoter and methods of use |
| WO2014164116A1 (en) | 2013-03-13 | 2014-10-09 | Pioneer Hi-Bred International, Inc. | Functional expression of bacterial major facilitator superfamily (sfm) gene in maize to improve agronomic traits and grain yield |
| WO2014159306A1 (en) | 2013-03-13 | 2014-10-02 | Pioneer Hi-Bred International, Inc. | Glyphosate application for weed control in brassica |
| WO2014160383A1 (en) | 2013-03-13 | 2014-10-02 | E. I. Dupont De Nemours & Company | Production of small interfering rnas in planta |
| WO2014164074A1 (en) | 2013-03-13 | 2014-10-09 | Pioneer Hi-Bred International, Inc. | Enhanced nitrate uptake and nitrate translocation by over-expressing maize functional low-affinity nitrate transporters in transgenic maize |
| EP3744727A1 (en) | 2013-03-14 | 2020-12-02 | Pioneer Hi-Bred International, Inc. | Compositions and methods to control insect pests |
| WO2014153234A1 (en) | 2013-03-14 | 2014-09-25 | Pioneer Hi-Bred International, Inc. | Compositions having dicamba decarboxylase activity and methods of use |
| WO2014160122A1 (en) | 2013-03-14 | 2014-10-02 | Pioneer Hi-Bred International, Inc. | Maize stress related transcription factor 18 and uses thereof |
| WO2014153254A2 (en) | 2013-03-14 | 2014-09-25 | Pioneer Hi-Bred International Inc. | Compositions and methods to control insect pests |
| WO2014153242A1 (en) | 2013-03-14 | 2014-09-25 | Pioneer Hi-Bred International, Inc. | Compositions having dicamba decarboxylase activity and methods of use |
| WO2014152507A2 (en) | 2013-03-15 | 2014-09-25 | Pioneer Hi-Bred International, Inc. | Modulation of acc deaminase expression |
| WO2014150914A2 (en) | 2013-03-15 | 2014-09-25 | Pioneer Hi-Bred International, Inc. | Phi-4 polypeptides and methods for their use |
| WO2014143996A2 (en) | 2013-03-15 | 2014-09-18 | Pioneer Hi-Bred International, Inc. | Compositions and methods of use of acc oxidase polynucleotides and polypeptides |
| WO2014201511A1 (en) | 2013-06-21 | 2014-12-24 | Gary David Housley | Method and apparatus for close-field electroporation |
| US11459579B2 (en) | 2013-07-09 | 2022-10-04 | Board Of Trustees Of Michigan State University | Transgenic plants produced with a K-domain, and methods and expression cassettes related thereto |
| US10570409B2 (en) | 2013-07-09 | 2020-02-25 | Board Of Trustees Of Michigan State University | Transgenic plants produced with a K-domain, and methods and expression cassettes related thereto |
| WO2015006105A1 (en) | 2013-07-09 | 2015-01-15 | Board Of Trustees Of Michigan State University | Transgenic plants produced with a k-domain, and methods and expression cassettes related thereto |
| WO2015017510A1 (en) | 2013-07-31 | 2015-02-05 | E. I. Du Pont De Nemours And Company | Modification of soybean seed composition to enhance feed, food and other industrial applications of soybean products |
| WO2015021139A2 (en) | 2013-08-08 | 2015-02-12 | Pioneer Hi-Bred International, Inc. | Insecticidal polypeptides having broad spectrum activity and uses thereof |
| WO2015023846A2 (en) | 2013-08-16 | 2015-02-19 | Pioneer Hi-Bred International, Inc. | Insecticidal proteins and methods for their use |
| WO2015038622A1 (en) | 2013-09-11 | 2015-03-19 | Pioneer Hi-Bred International, Inc. | Plant regulatory elements and methods of use thereof |
| EP3692786A1 (en) | 2013-09-13 | 2020-08-12 | Pioneer Hi-Bred International, Inc. | Insecticidal proteins and methods for their use |
| EP4159028A1 (en) | 2013-09-13 | 2023-04-05 | Pioneer Hi-Bred International, Inc. | Insecticidal proteins and methods for their use |
| WO2015038734A2 (en) | 2013-09-13 | 2015-03-19 | Pioneer Hi-Bred International, Inc. | Insecticidal proteins and methods for their use |
| US10428336B2 (en) | 2013-10-16 | 2019-10-01 | The Australian National University | Method for modulating plant growth |
| WO2015057600A1 (en) | 2013-10-18 | 2015-04-23 | E. I. Du Pont De Nemours And Company | Glyphosate-n-acetyltransferase (glyat) sequences and methods of use |
| WO2015066011A2 (en) | 2013-10-29 | 2015-05-07 | Pioneer Hi-Bred International, Inc. | Self-reproducing hybrid plants |
| WO2015095804A1 (en) | 2013-12-19 | 2015-06-25 | Amyris, Inc. | Methods for genomic integration |
| EP3705489A1 (en) | 2014-02-07 | 2020-09-09 | Pioneer Hi-Bred International, Inc. | Insecticidal proteins and methods for their use |
| WO2015120276A1 (en) | 2014-02-07 | 2015-08-13 | Pioneer Hi Bred International Inc | Insecticidal proteins and methods for their use |
| WO2015120270A1 (en) | 2014-02-07 | 2015-08-13 | Pioneer Hi Bred International, Inc. | Insecticidal proteins and methods for their use |
| WO2015164457A1 (en) | 2014-04-22 | 2015-10-29 | E. I. Du Pont De Nemours And Company | Plastidic carbonic anhydrase genes for oil augmentation in seeds with increased dgat expression |
| WO2016000647A1 (en) | 2014-07-03 | 2016-01-07 | Pioneer Overseas Corporation | Plants having enhanced tolerance to insect pests and related constructs and methods involving insect tolerance genes |
| WO2016022516A1 (en) | 2014-08-08 | 2016-02-11 | Pioneer Hi-Bred International, Inc. | Ubiquitin promoters and introns and methods of use |
| EP3628739A1 (en) | 2014-09-12 | 2020-04-01 | E. I. du Pont de Nemours and Company | Generation of site-specific-integration sites for complex trait loci in corn and soybean, and methods of use |
| US11560568B2 (en) | 2014-09-12 | 2023-01-24 | E. I. Du Pont De Nemours And Company | Generation of site-specific-integration sites for complex trait loci in corn and soybean, and methods of use |
| WO2016040030A1 (en) | 2014-09-12 | 2016-03-17 | E. I. Du Pont De Nemours And Company | Generation of site-specific-integration sites for complex trait loci in corn and soybean, and methods of use |
| WO2016044092A1 (en) | 2014-09-17 | 2016-03-24 | Pioneer Hi Bred International Inc | Compositions and methods to control insect pests |
| WO2016061206A1 (en) | 2014-10-16 | 2016-04-21 | Pioneer Hi-Bred International, Inc. | Insecticidal proteins and methods for their use |
| WO2016061197A1 (en) | 2014-10-16 | 2016-04-21 | Pioneer Hi-Bred International, Inc. | Insecticidal polypeptides having improved activity spectrum and uses thereof |
| WO2016060949A1 (en) | 2014-10-16 | 2016-04-21 | Pioneer Hi-Bred International, Inc. | Insecticidal polypeptides having broad spectrum activity and uses thereof |
| WO2016094165A1 (en) | 2014-12-12 | 2016-06-16 | Syngenta Participations Ag | Compositions and methods for controlling plant pests |
| EP3795582A1 (en) | 2014-12-12 | 2021-03-24 | Syngenta Participations Ag | Compositions and methods for controlling plant pests |
| WO2016100333A1 (en) | 2014-12-15 | 2016-06-23 | Syngenta Participations Ag | Pesticidal microrna carriers and use thereof |
| WO2016100309A1 (en) | 2014-12-16 | 2016-06-23 | Pioneer Hi-Bred International, Inc. | Restoration of male fertility in wheat |
| WO2016099916A1 (en) | 2014-12-19 | 2016-06-23 | E. I. Du Pont De Nemours And Company | Polylactic acid compositions with accelerated degradation rate and increased heat stability |
| WO2016105696A1 (en) | 2014-12-23 | 2016-06-30 | Syngenta Participations Ag | Biological control of coleopteran pests |
| WO2016144688A1 (en) | 2015-03-11 | 2016-09-15 | Pioneer Hi Bred International Inc | Insecticidal combinations of pip-72 and methods of use |
| WO2016149352A1 (en) | 2015-03-19 | 2016-09-22 | Pioneer Hi-Bred International Inc | Methods and compositions for accelerated trait introgression |
| EP4118955A1 (en) | 2015-03-19 | 2023-01-18 | Pioneer Hi-Bred International, Inc. | Compositions for accelerated trait introgression |
| WO2016177887A1 (en) | 2015-05-07 | 2016-11-10 | Limagrain Europe | Polynucleotide responsible of haploid induction in maize plants and related processes |
| EP3091076A1 (en) | 2015-05-07 | 2016-11-09 | Limagrain Europe | Polynucleotide responsible of haploid induction in maize plants and related processes |
| EP3453764A1 (en) | 2015-05-07 | 2019-03-13 | Institut National De La Recherche Agronomique (INRA) | Polynucleotide responsible of haploid induction in maize plants and related processes |
| WO2016186986A1 (en) | 2015-05-19 | 2016-11-24 | Pioneer Hi Bred International Inc | Insecticidal proteins and methods for their use |
| WO2016205445A1 (en) | 2015-06-16 | 2016-12-22 | Pioneer Hi-Bred International, Inc. | Compositions and methods to control insect pests |
| WO2017023486A1 (en) | 2015-08-06 | 2017-02-09 | Pioneer Hi-Bred International, Inc. | Plant derived insecticidal proteins and methods for their use |
| EP3943602A1 (en) | 2015-08-06 | 2022-01-26 | Pioneer Hi-Bred International, Inc. | Plant derived insecticidal proteins and methods for their use |
| WO2017040343A1 (en) | 2015-08-28 | 2017-03-09 | Pioneer Hi-Bred International, Inc. | Ochrobactrum-mediated transformation of plants |
| WO2017066597A1 (en) | 2015-10-16 | 2017-04-20 | Pioneer Hi-Bred International, Inc. | Generating maize plants with enhanced resistance to northern leaf blight |
| WO2017074547A1 (en) | 2015-10-30 | 2017-05-04 | Pioneer Hi-Bred International, Inc. | Methods and compositions for rapid plant transformation |
| WO2017079026A1 (en) | 2015-11-06 | 2017-05-11 | E. I. Du Pont De Nemours And Company | Generation of complex trait loci in soybean and methods of use |
| WO2017078836A1 (en) | 2015-11-06 | 2017-05-11 | Pioneer Hi-Bred International, Inc. | Methods and compositions of improved plant transformation |
| WO2017112006A1 (en) | 2015-12-22 | 2017-06-29 | Pioneer Hi-Bred International, Inc. | Tissue-preferred promoters and methods of use |
| EP4257694A2 (en) | 2015-12-22 | 2023-10-11 | Pioneer Hi-Bred International, Inc. | Tissue-preferred promoters and methods of use |
| WO2017161264A1 (en) | 2016-03-18 | 2017-09-21 | Pioneer Hi-Bred International, Inc. | Methods and compositions for producing clonal, non-reduced, non-recombined gametes |
| WO2017180715A2 (en) | 2016-04-14 | 2017-10-19 | Pioneer Hi-Bred International, Inc. | Insecticidal polypeptides having improved activity spectrum and uses thereof |
| WO2017184673A1 (en) | 2016-04-19 | 2017-10-26 | Pioneer Hi-Bred International, Inc. | Insecticidal combinations of polypeptides having improved activity spectrum and uses thereof |
| WO2017192560A1 (en) | 2016-05-04 | 2017-11-09 | Pioneer Hi-Bred International, Inc. | Insecticidal proteins and methods for their use |
| EP3960863A1 (en) | 2016-05-04 | 2022-03-02 | Pioneer Hi-Bred International, Inc. | Insecticidal proteins and methods for their use |
| EP4219731A2 (en) | 2016-05-18 | 2023-08-02 | Amyris, Inc. | Compositions and methods for genomic integration of nucleic acids into exogenous landing pads |
| WO2017218207A1 (en) | 2016-06-16 | 2017-12-21 | Pioneer Hi-Bred International, Inc. | Compositions and methods to control insect pests |
| EP4083215A1 (en) | 2016-06-24 | 2022-11-02 | Pioneer Hi-Bred International, Inc. | Plant regulatory elements and methods of use thereof |
| WO2017222821A2 (en) | 2016-06-24 | 2017-12-28 | Pioneer Hi-Bred International, Inc. | Plant regulatory elements and methods of use thereof |
| WO2018005411A1 (en) | 2016-07-01 | 2018-01-04 | Pioneer Hi-Bred International, Inc. | Insecticidal proteins from plants and methods for their use |
| EP3954202A1 (en) | 2016-07-01 | 2022-02-16 | Pioneer Hi-Bred International, Inc. | Insecticidal proteins from plants and methods for their use |
| WO2018013333A1 (en) | 2016-07-12 | 2018-01-18 | Pioneer Hi-Bred International, Inc. | Compositions and methods to control insect pests |
| WO2018026774A1 (en) | 2016-08-05 | 2018-02-08 | Syngenta Participations Ag | Control of coleopteran pests using rna molecules |
| EP4079857A2 (en) | 2016-08-05 | 2022-10-26 | Syngenta Participations Ag | Control of coleopteran pests using rna molecules |
| US11140902B2 (en) | 2016-09-27 | 2021-10-12 | University Of Florida Research Foundation, Inc. | Insect toxin delivery mediated by a densovirus coat protein |
| WO2018078390A1 (en) | 2016-10-31 | 2018-05-03 | Institute Of Genetics And Developmental Biology Chinese Academy Of Sciences | Compositions and methods for enhancing abiotic stress tolerance |
| WO2018084936A1 (en) | 2016-11-01 | 2018-05-11 | Pioneer Hi-Bred International, Inc. | Insecticidal proteins and methods for their use |
| EP4050021A1 (en) | 2016-11-01 | 2022-08-31 | Pioneer Hi-Bred International, Inc. | Insecticidal proteins and methods for their use |
| WO2018111551A1 (en) | 2016-12-14 | 2018-06-21 | Pioneer Hi-Bred International, Inc. | Insecticidal proteins and methods for their use |
| WO2018118811A1 (en) | 2016-12-22 | 2018-06-28 | Pioneer Hi-Bred International, Inc. | Insecticidal proteins and methods for their use |
| WO2018140214A1 (en) | 2017-01-24 | 2018-08-02 | Pioneer Hi-Bred International, Inc. | Nematicidal protein from pseudomonas |
| WO2018148001A1 (en) | 2017-02-08 | 2018-08-16 | Pioneer Hi-Bred International Inc | Insecticidal combinations of plant derived insecticidal proteins and methods for their use |
| WO2018208882A1 (en) | 2017-05-11 | 2018-11-15 | Pioneer Hi-Bred International, Inc. | Insecticidal proteins and methods for their use |
| WO2018217333A1 (en) | 2017-05-26 | 2018-11-29 | Pioneer Hi-Bred International, Inc. | Insecticidal polypeptides having improved activity spectrum and uses thereof |
| WO2019049111A1 (en) | 2017-09-11 | 2019-03-14 | R. J. Reynolds Tobacco Company | Methods and compositions for increasing expression of genes of interest in a plant by co-expression with p21 |
| WO2019060383A1 (en) | 2017-09-25 | 2019-03-28 | Pioneer Hi-Bred, International, Inc. | Tissue-preferred promoters and methods of use |
| WO2019074598A1 (en) | 2017-10-13 | 2019-04-18 | Pioneer Hi-Bred International, Inc. | Virus-induced gene silencing technology for insect control in maize |
| WO2019076891A2 (en) | 2017-10-18 | 2019-04-25 | Syngenta Participations Ag | Control of hemipteran pests using rna molecules |
| EP4122947A1 (en) | 2017-12-19 | 2023-01-25 | Pioneer Hi-Bred International, Inc. | Insecticidal polypeptides and uses thereof |
| US11905518B2 (en) | 2018-02-12 | 2024-02-20 | Curators Of The University Of Missouri | Small auxin upregulated (SAUR) gene for the improvement of root system architecture, waterlogging tolerance, drought resistance and yield in plants and methods of uses |
| WO2019162163A1 (en) | 2018-02-26 | 2019-08-29 | Devgen Nv | Control of insect pests using rna molecules |
| WO2019169150A1 (en) | 2018-03-02 | 2019-09-06 | Pioneer Hi-Bred International, Inc. | Plant health assay |
| WO2019206780A1 (en) | 2018-04-27 | 2019-10-31 | Devgen Nv | Control of insect pests using rna molecules |
| WO2019226508A1 (en) | 2018-05-22 | 2019-11-28 | Pioneer Hi-Bred International, Inc. | Plant regulatory elements and methods of use thereof |
| WO2020005933A1 (en) | 2018-06-28 | 2020-01-02 | Pioneer Hi-Bred International, Inc. | Methods for selecting transformed plants |
| WO2020046701A1 (en) | 2018-08-29 | 2020-03-05 | Pioneer Hi-Bred International, Inc. | Insecticidal proteins and methods for their use |
| WO2020092487A1 (en) | 2018-10-31 | 2020-05-07 | Pioneer Hi-Bred International, Inc. | Compositions and methods for ochrobactrum-mediated plant transformation |
| WO2020185751A1 (en) | 2019-03-11 | 2020-09-17 | Pioneer Hi-Bred International, Inc. | Methods for clonal plant production |
| WO2020187798A1 (en) | 2019-03-21 | 2020-09-24 | Devgen Nv | Control of insect pests using rna molecules |
| WO2020198408A1 (en) | 2019-03-27 | 2020-10-01 | Pioneer Hi-Bred International, Inc. | Plant explant transformation |
| WO2020198496A1 (en) | 2019-03-28 | 2020-10-01 | Pioneer Hi-Bred International, Inc. | Modified agrobacterium strains and use thereof for plant transformation |
| WO2020214986A1 (en) | 2019-04-18 | 2020-10-22 | Pioneer Hi-Bred International, Inc. | Embryogenesis factors for cellular reprogramming of a plant cell |
| EP4331364A2 (en) | 2020-02-04 | 2024-03-06 | Corteva Agriscience LLC | Compositions having pesticidal utility and processes related thereto |
| WO2021158455A1 (en) | 2020-02-04 | 2021-08-12 | Dow Agrosciences Llc | Compositions having pesticidal utility and processes related thereto |
| WO2021252238A1 (en) | 2020-06-12 | 2021-12-16 | Pioneer Hi-Bred International, Inc. | Alteration of seed composition in plants |
| WO2021257206A1 (en) | 2020-06-17 | 2021-12-23 | Pioneer Hi-Bred International, Inc. | Generating maize plants with enhanced resistance to northern leaf blight |
| WO2022015619A2 (en) | 2020-07-14 | 2022-01-20 | Pioneer Hi-Bred International, Inc. | Insecticidal proteins and methods for their use |
| WO2022035649A1 (en) | 2020-08-10 | 2022-02-17 | Pioneer Hi-Bred International, Inc. | Plant regulatory elements and methods of use thereof |
| WO2022035653A2 (en) | 2020-08-10 | 2022-02-17 | E. I. Du Pont De Nemours And Company | Compositions and methods for enhancing resistance to northern leaf blight in maize |
| WO2022072335A2 (en) | 2020-09-30 | 2022-04-07 | Pioneer Hi-Bred International, Inc. | Rapid transformation of monocot leaf explants |
| WO2022087616A1 (en) | 2020-10-21 | 2022-04-28 | Pioneer Hi-Bred International, Inc. | Parthenogenesis factors and methods of using same |
| WO2022115524A2 (en) | 2020-11-24 | 2022-06-02 | AgBiome, Inc. | Pesticidal genes and methods of use |
| WO2022226316A1 (en) | 2021-04-22 | 2022-10-27 | Precision Biosciences, Inc. | Compositions and methods for generating male sterile plants |
| WO2022236060A1 (en) | 2021-05-06 | 2022-11-10 | AgBiome, Inc. | Pesticidal genes and methods of use |
| US11895964B2 (en) | 2021-06-02 | 2024-02-13 | Nutrien Ag Solutions, Inc. | Inbred rice line DG263L |
| WO2023012342A1 (en) | 2021-08-06 | 2023-02-09 | Kws Vegetables B.V. | Durable downy mildew resistance in spinach |
| WO2023107943A1 (en) | 2021-12-07 | 2023-06-15 | AgBiome, Inc. | Pesticidal genes and methods of use |
| WO2023183918A1 (en) | 2022-03-25 | 2023-09-28 | Pioneer Hi-Bred International, Inc. | Methods of parthenogenic haploid induction and haploid chromosome doubling |
| WO2023224815A2 (en) | 2022-05-18 | 2023-11-23 | Corteva Agriscience Llc | Compositions having pesticidal utility and processes related thereto |
| WO2024023578A1 (en) | 2022-07-28 | 2024-02-01 | Institut Pasteur | Hsc70-4 in host-induced and spray-induced gene silencing |
| WO2024044596A1 (en) | 2022-08-23 | 2024-02-29 | AgBiome, Inc. | Pesticidal genes and methods of use |
Also Published As
Similar Documents
| Publication | Publication Date | Title |
|---|---|---|
| AU760113C (en) | Compositions and methods for genetic modification of plants | |
| US7462766B2 (en) | Compositions comprising non-identical recombination sites | |
| US20020132350A1 (en) | Targeted genetic manipulation using Mu bacteriophage cleaved donor complex | |
| EP2612918A1 (en) | In planta recombination | |
| AU2006203210B2 (en) | Compositions and methods for genetic modification of plants | |
| WO2003062425A1 (en) | Short fragment homologous recombination to effect targeted genetic alterations in plants | |
| AU1157399A (en) | Methods for obtaining plant varieties |
Legal Events
| Date | Code | Title | Description |
|---|---|---|---|
| AK | Designated states |
Kind code of ref document: A1 Designated state(s): AL AM AT AT AU AZ BA BB BG BR BY CA CH CN CU CZ CZ DE DE DK DK EE EE ES FI FI GB GE GH GM HR HU ID IL IS JP KE KG KP KR KZ LC LK LR LS LT LU LV MD MG MK MN MW MX NO NZ PL PT RO RU SD SE SG SI SK SK SL TJ TM TR TT UA UG US UZ VN YU ZW |
|
| AL | Designated countries for regional patents |
Kind code of ref document: A1 Designated state(s): GH GM KE LS MW SD SZ UG ZW AM AZ BY KG KZ MD RU TJ TM AT BE CH CY DE DK ES FI FR GB GR IE IT LU MC NL PT SE BF BJ CF CG CI CM GA GN GW ML MR NE SN TD TG |
|
| 121 | Ep: the epo has been informed by wipo that ep was designated in this application | ||
| DFPE | Request for preliminary examination filed prior to expiration of 19th month from priority date (pct application filed before 20040101) | ||
| WWE | Wipo information: entry into national phase |
Ref document number: 503859 Country of ref document: NZ |
|
| ENP | Entry into the national phase |
Ref document number: 2306184 Country of ref document: CA Ref country code: CA Ref document number: 2306184 Kind code of ref document: A Format of ref document f/p: F |
|
| WWE | Wipo information: entry into national phase |
Ref document number: 14629/99 Country of ref document: AU |
|
| NENP | Non-entry into the national phase |
Ref country code: KR |
|
| WWE | Wipo information: entry into national phase |
Ref document number: 1998958629 Country of ref document: EP |
|
| WWP | Wipo information: published in national office |
Ref document number: 1998958629 Country of ref document: EP |
|
| REG | Reference to national code |
Ref country code: DE Ref legal event code: 8642 |
|
| WWG | Wipo information: grant in national office |
Ref document number: 14629/99 Country of ref document: AU |
|
| WWG | Wipo information: grant in national office |
Ref document number: 1998958629 Country of ref document: EP |